
原标题:【地球演义】第二百二十回 嘎啦簸箕翻身记
“嘎啦簸箕”是井蛙老家对双壳类软体动物(其实老家有的也就是河蚌河蚬一类的淡水双壳类)的俗称。“嘎啦”应该是“蛤蜊”的变音,“簸箕”应该是形容贝壳的形状。井蛙媳妇说她们家乡话里,双壳类叫“歪(三声)歪(轻声)”,一下子又萌了起来。人民群众的语言真是非常生动啊。
上回介绍了古生代腕足动物的兴衰,这一回就来聊聊它们的老对手双壳类软体动物。软体动物门基本上和腕足动物门(托莫特壳类)在寒武纪早期同时出现,不过最早的软体动物应该是一些用腹部爬行,吞食底泥或者啃食藻类的种类。演化出双壳类这种潜居滤食的类型要稍微迟一点。

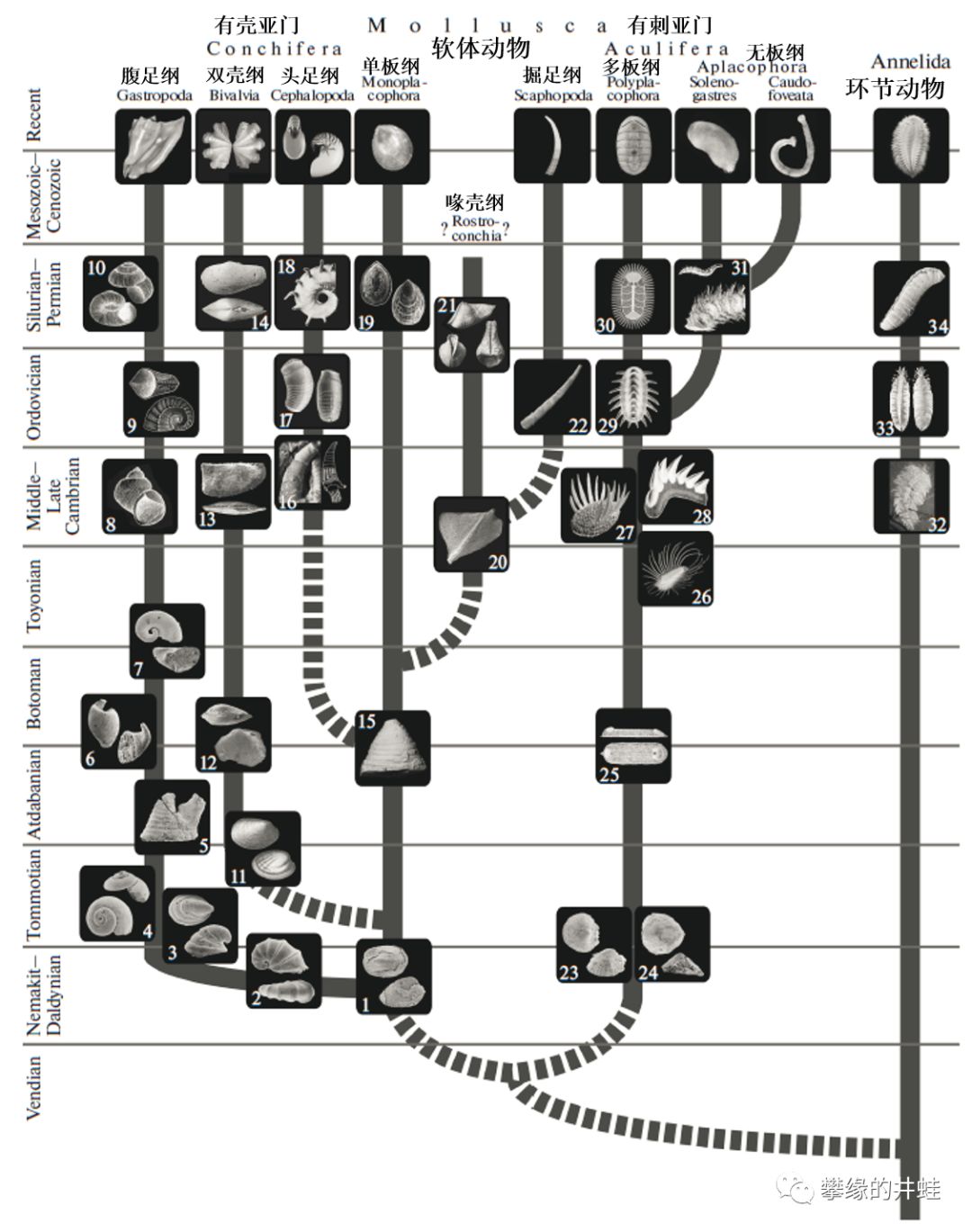
软体动物主要类群和早期演化的代表种类。除了喙壳纲在二叠纪折戟之外,其他几大类都成功延续到现代,这实在是一项非常了不起的成就呢。图片来源自[1]。
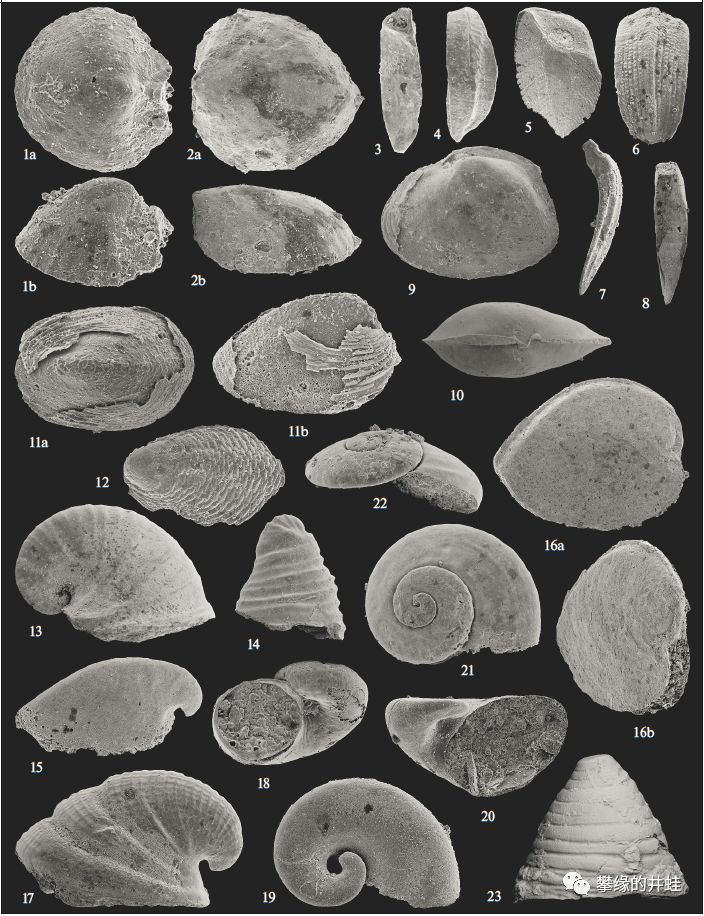
一些早寒武世的软体动物化石代表。1–8:多板纲的壳板;9,10:双壳纲的内模;11,12,23:单板纲;13–22:腹足纲。这些贝壳都在1毫米左右,当时的软体动物全都非常小,但基本形态已已经和今天相差无几了,一眼就可以辨认出来。图片来源自[1]。
也许就是迟来的这一步,让腕足动物占得先机,遍地开花。在之后的3亿年里,腕足动物一直都占据着“贝类”的正统地位,把高度趋(wu)同(chi)演(chao)化(xi)的双壳类按在地上摩擦。
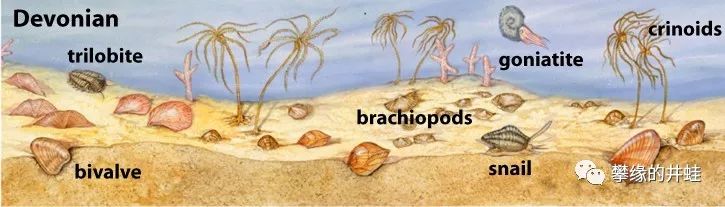
泥盆纪的海洋底栖无脊椎动物。腕足动物的种类和数量远超双壳类。图片来源自网络。
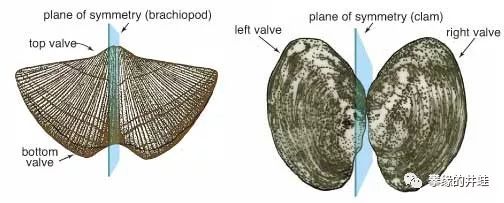
腕足动物(左)和双壳类(右)是如此相似,以至于很多人都弄不清二者的区别。其实,腕足动物的两片贝壳是背壳和腹壳(可以想象一下乌龟的背甲和腹甲),所以两片贝壳形状不同,但每片贝壳都左右对称(当然有例外)。而双壳类的贝壳是左右两片,两片贝壳互相对称(当然也有例外)。图片来源自网络。
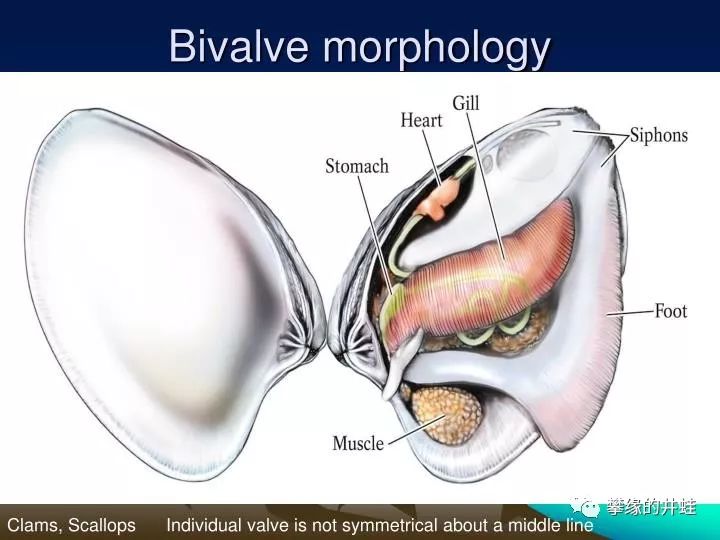
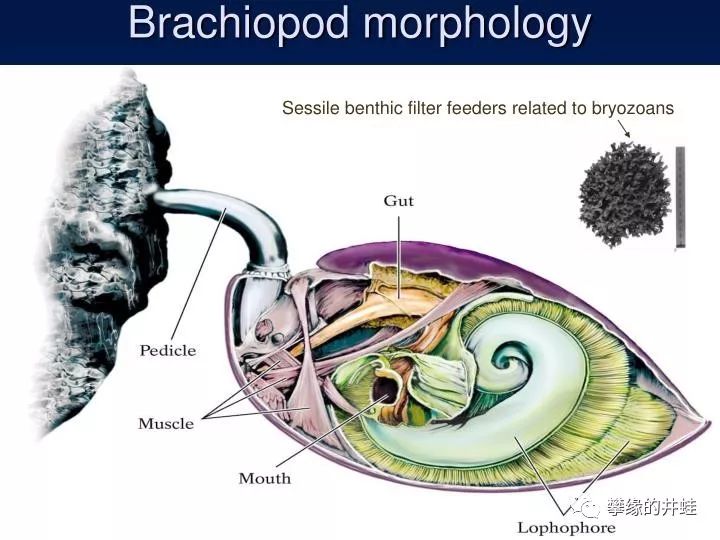
从内部结构上看,二者都演化出贝壳之间的铰合结构,控制开闭的肌肉群,还有滤食的筛状器官。不过区别也很明显,双壳类有发达的斧足,可以从两壳中间伸出来,负责爬行和挖洞。而腕足动物只有一根棒状的肉茎。另外,双壳类的瓣鳃和腕足动物的腕螺也大不相同。图片来源见水印。
绝大多数腕足动物都不擅长在沉积物中挖掘洞穴(这可能和背-腹壳的结构和肉茎的生长位置有关)。它们要么附着在硬质的岩礁上,要么平躺在海床上。于是双壳类凭借天生的优势,在海底淤泥和沿岸滩涂立定了脚跟,平稳发展。

腕足动物中,只有舌形贝纲的少数种类在淤泥中掘穴生活。图片来源见水印。

泥涂是双壳类的龙兴之地,掘穴是双壳类的看家本领。从烂泥里扒出各种蛏蚌蛤蚬蚶,赶过海的朋友都知道这是多么快乐的体验。图片来源自网络。
双壳类的斧足强壮,灵活,可大可小,可粗可细,可软可硬,可进可退,而且布满粘液,是掘穴潜沙的利器。腕足动物的肉茎就没有这么强大的功能。图片来源自网络。
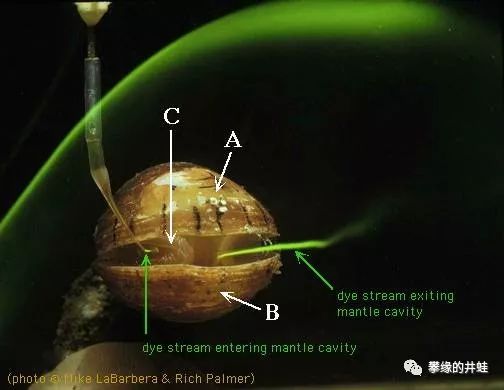
为适应在淤泥里摸爬滚打的生活,双壳类还练就了另一项绝技:进出水管(Siphon)系统。通过有规律地拍打外套膜和瓣鳃上的无数细小纤毛,在外套腔内形成压差,引导水流从进水口(Incurrent siphon)流入,穿过丛密的鳃丝,滤取其中的有机碎屑,吸收溶解氧,排出二氧化碳,兜转一圈后,再从出水口(Excurrent siphon)流出体外,顺便把肛门(开口在出水口附近)排出的废物冲走。


双壳类种类繁多,水管的形态差异极大,有些只是两个简单的开口(上图),也有的被发达的肌肉包裹,延伸成粗壮的肉柱(下图),但功能都是一样的。图片来源自网络。
可不要小看这套导流系统,它的出现使双壳类和腕足动物这两大底栖滤食类群在装备层面真正拉开了差距。
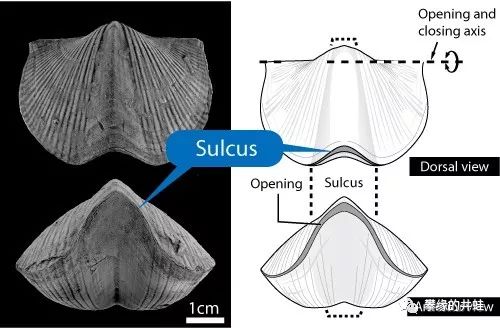
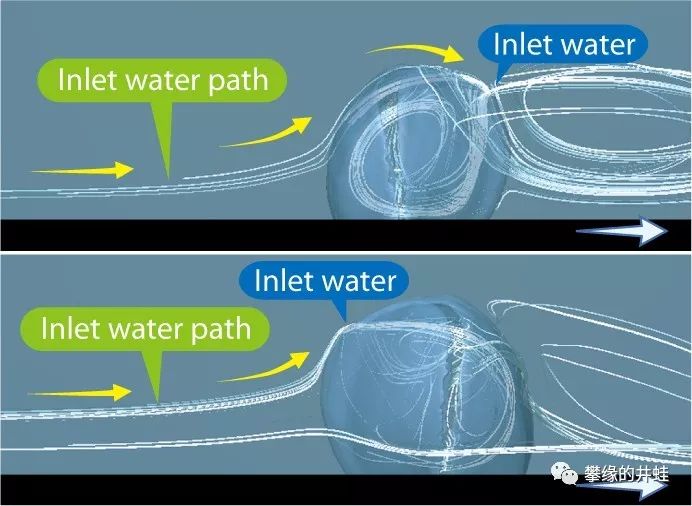
研究者模拟了水流经过泥盆纪腕足动物拟石燕Paraspirifer(上图化石)的贝壳时产生的湍流。拟石燕只需要稍微调整躺卧的角度和贝壳张开的大小,就可以让流水进入贝壳内部,流过触手冠,获得食物和溶解氧。图片来源自[2]。
另一种泥盆纪腕足动物弓石燕Cyrtospirifer(上图化石)的湍流模拟分析。图片来源自[3]。
必须承认,腕足动物的流体力学设计也是极其精巧的,仔细观察上面的模拟中,水流在贝壳内部的流向,和腕足动物的腕螺结构高度匹配,这样无疑可以增大接触面积,提高呼吸和滤食的效率。
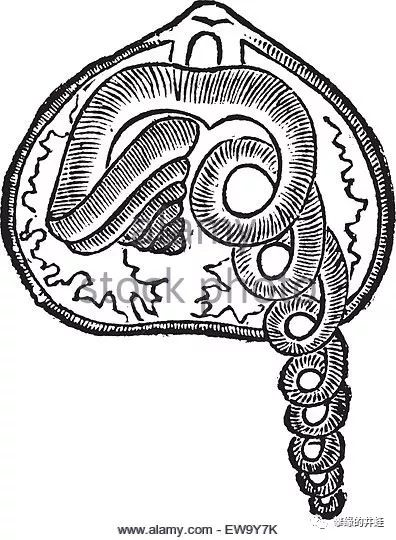
腕足动物的腕螺结构,和双壳类的瓣鳃完全不同,配合贝壳激起的湍流,是非常高效的滤食工具。图片来源见水印。
但是无论怎样,腕足动物属于“被动滤食者(Passive Filter-Feeder)”,只能被动地接受水流带来的悬浮颗粒。水流的速度,方向,有机物携带量都会制约腕足动物的生存。而双壳类被归为“主动滤食者(Active Filter-Feeder)”,它们可以自己制造水流,激荡起水下的沉积物,源源不断地获取食物。加上双壳类具有斧足,虽然缓慢,但也足以自由移动,寻觅松软富饶的栖息地。所以双壳类比腕足动物摄食能力更强,它们可以适应水流平缓,和有机悬浮物较少的水体。这也许就是双壳类能够成功进入淡水,而腕足动物却不行的原因吧。

进出水系统的功能非常强大,足以让双壳类获得足够的营养,演化出砗磲那样的巨无霸(当然在古生代,还有更加巨大的双壳类,到时候再介绍)。这个体型是腕足动物,比如大长身贝之流望尘莫及的。图片来源见水印。
水流通过一种现代腕足动物腕螺的流向。现代淡水生的底栖滤食动物,比如水螅虫和苔藓虫都是一些非常微小的个体。唯有双壳类能够长到相当大的体型,这也可以证明主动滤食的高效。图片来源见水印。
因为缺乏软体结构的化石,很难考证双壳类到底是在什么时候演化出进出水系统的。不过到古生代晚期,双壳类已经成功进入淡水,体型明显增大,种类和数量也显著增加,你甚至可以从中找到许多熟悉的身影。
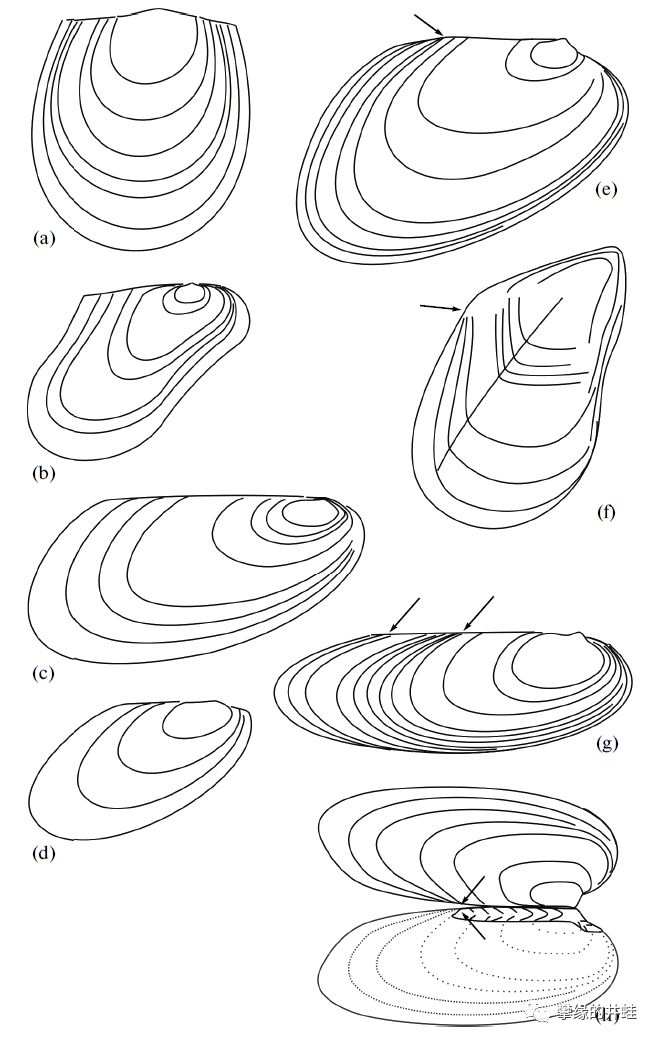
一些二叠纪双壳类的外形和生长方式。多样性已经很高了。图片来源自[4]。

巴西帕拉娜盆地(Paraná Basin)发现的二叠纪海洋双壳类化石。A-F:Aviculopectenmultiscalptus(燕海扇科);G-M:Myonia argentinensis(Megadesmidae科)。图片来源自[5],标尺长度1厘米。

这是一种现代海扇科双壳类。图片来源自网络。


一些东欧地区发现的二叠纪淡水双壳类化石,外形很像今天的河蚌。双壳类在石炭纪成功入侵淡水,到二叠纪已经遍布泛古陆的各大江河湖泊,一直延续到今天。而它们的老对手腕足动物从没能跨出海洋一步。图片来源自[6]。

这是一种现代河蚌目双壳类(其中混杂了一些其他动植物)。图片来源自网络。
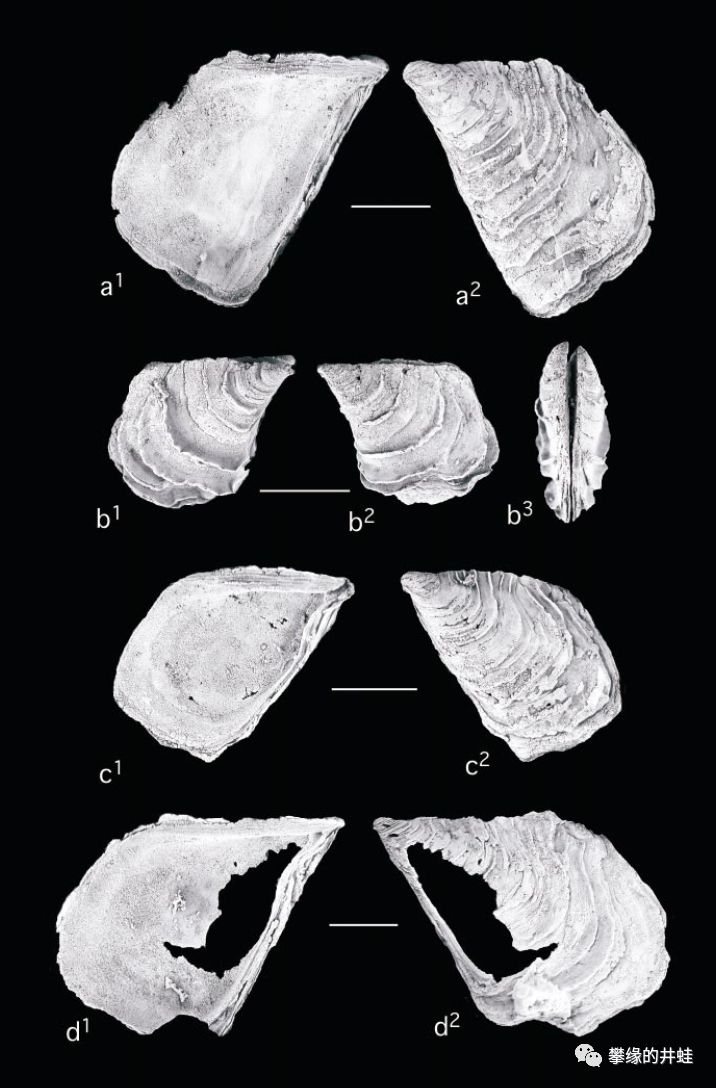
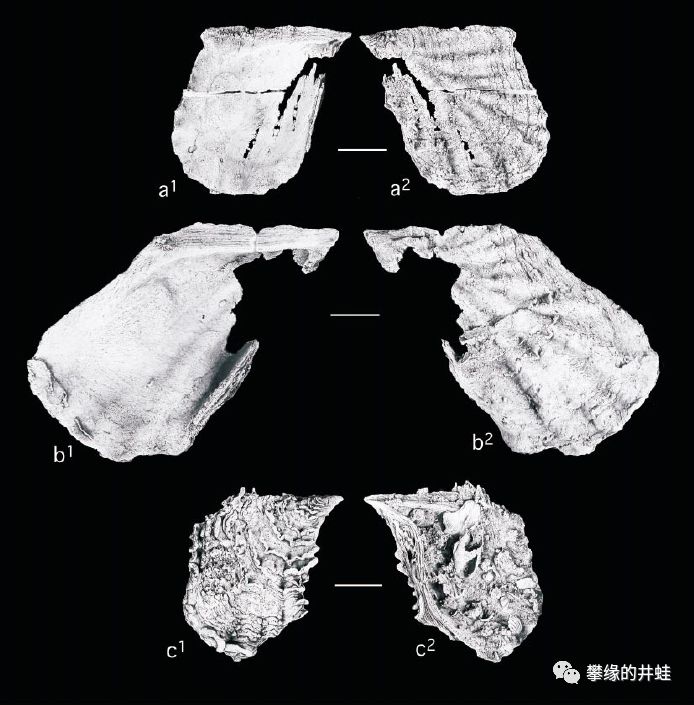
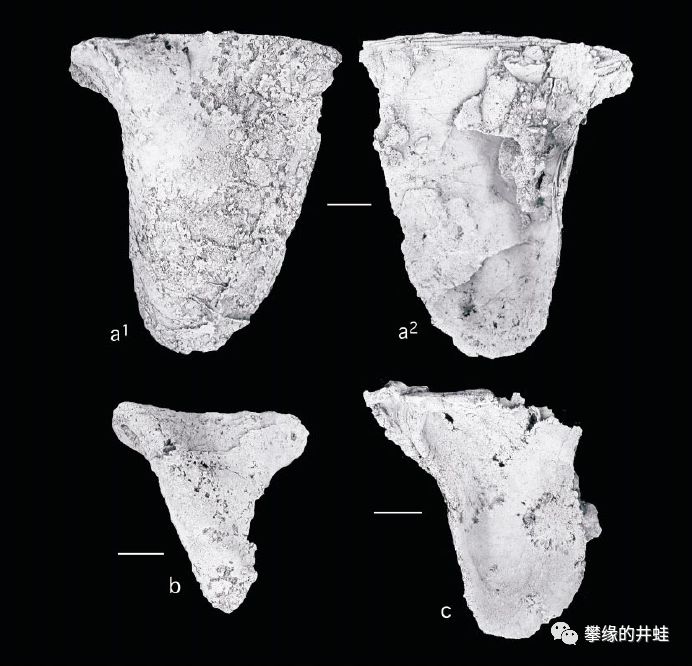
一些德克萨斯发现的二叠纪海洋双壳类化石。上:Myalina lamellosa;中:Myalinaplicata;下:Myalina copei。它们属于已灭绝的Myalinidae科,是莺蛤目(Pterioida)的早期种类。图片来源自[7],标尺长度1厘米。

这是一种现代莺蛤目双壳类(江珧科)。它们的贝壳形状非常接近,很可能具有相似的味道,不是,生活习性。图片来源自网络。
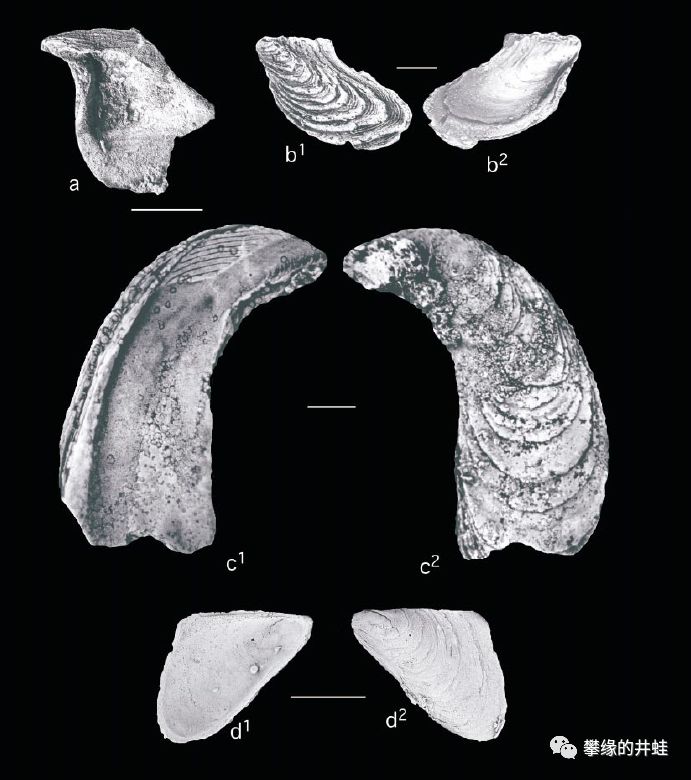
另一些德克萨斯发现的二叠纪海洋双壳类化石。a,b:Elversella rugosa;c:Novaculapermia boydi;d:Myalinella acutirostris。图片来源自[7],标尺长度1厘米。
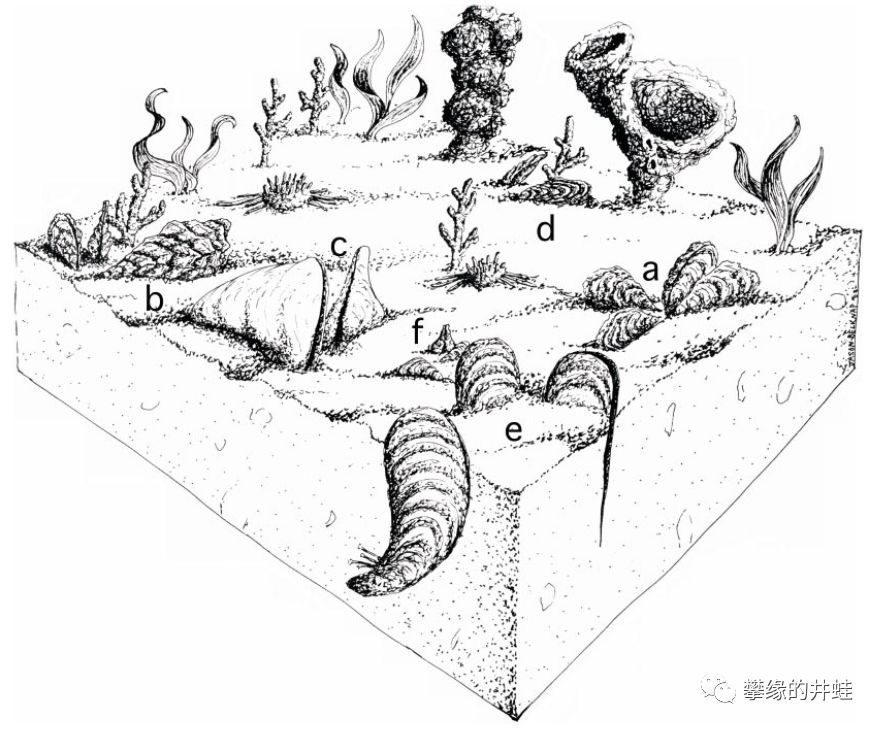
二叠纪海底一角的复原图。各种双壳类潜穴而居,就像今天的浅海滩涂一样。a:Myalina lamellosa;b:Myalina plicata;c:Myalina copei;d:Elversella rugosa;e:Novaculapermia boydi;f:Myalinella acutirostris。图片来源自[7]。
一些二叠纪的双壳类甚至开始离开涂泥,去侵占腕足动物的领地——岩礁。这可是个不寻常的信号。

在德克萨斯玻璃山地区(Glass Mountains)发现了最早的附礁双壳类化石——二叠纪的Prospondylus acinetus。这种软体动物用右侧贝壳(A)把自己牢牢粘贴在礁岩上,张开左侧贝壳(B)过滤食物,就像今天的牡蛎那样。图片来源自[8]。

说到牡蛎,我就想起了。。。图片来源自网络。
至此,双壳类的装备水平和适应能力已经超越了腕足动物,甚至已经在局部发起了反攻。腕足动物所剩的,只有先发优势带来的庞大体量了。其实,底栖滤食动物之间的竞争并不算太激烈,双壳类要挤掉腕足动物,本来会是一个很缓慢的过程。然而二叠纪末的大灭绝事件几乎对地球上的所有生物发动了无差别攻击,无论双壳类还是腕足动物全都损失惨重,二者又被硬生生地拉回同一条起跑线上。当灾难过去,地球生态系统开始恢复时,腕足动物的先发优势已经荡然无存,于是双壳类在3亿年的漫长等待后,终于迎来了压倒性的胜利。
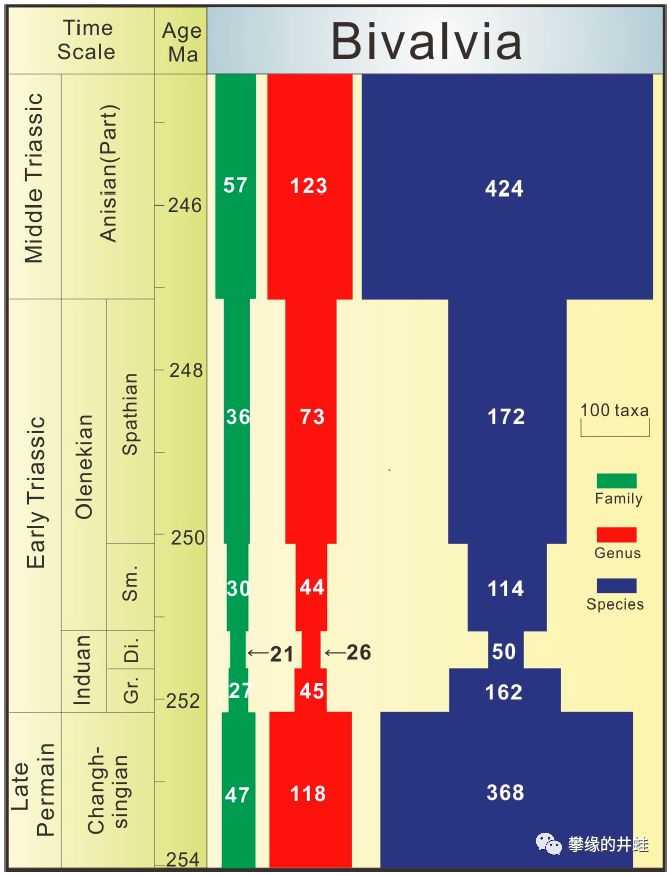
双壳类在二叠纪末大灭绝中受到的冲击和在三叠纪早期的恢复。绿色柱为科数,红色柱为属数,蓝色柱为种类数。经过400万年的复苏,双壳类的多样性已经赶上并超过了大灭绝发生前的水平。而腕足动物,上回介绍过,再也没能恢复元气。图片来源自[9]。
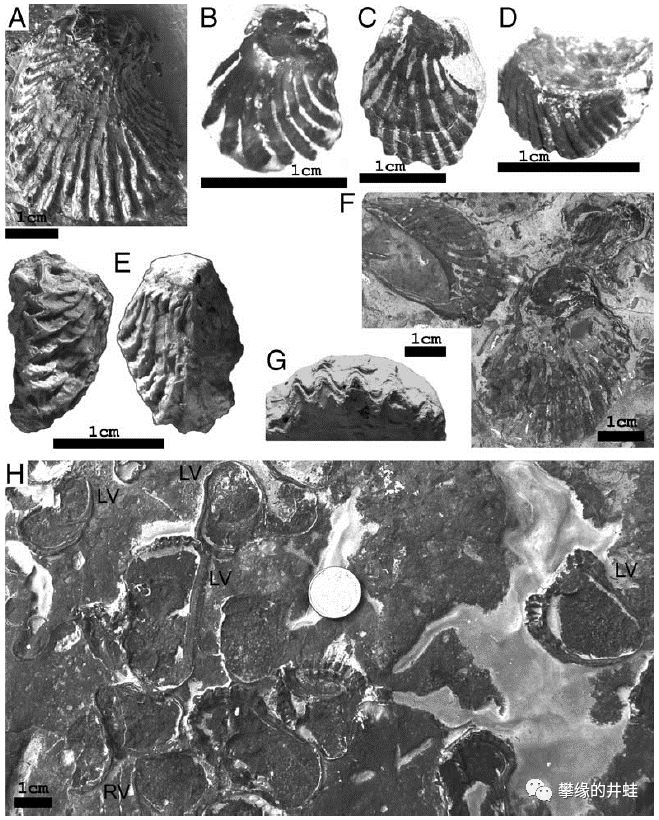
附礁生活的双壳类在大灭绝中幸存下来,由于宿敌腕足动物的衰退,它们越发兴盛,迅速演化出丰富的种类,占据了更加广阔的岩礁。上图是一些德国发现的三叠纪附礁双壳类的化石。A:一只Umbrostrea spondyloides附着在Umbrostreacristadifformis的贝壳上;B-E,G:Umbrostrea cristadifformis;F,H:Umbrostrea spondyloides。图片来源自[8]。
双壳类兴起的意义可能远比我们想象的要大得多。和腕足动物相比,双壳类分布更广,体型更大,营养更丰富。作为食物链的重要环节,催生出各种以蛤贝为食的新兴动物类群,比如三叠纪崛起的海洋爬行类,侏罗纪出现的十足目虾蟹和鳐鱼等软骨鱼类。海洋生态系统被重新塑造。
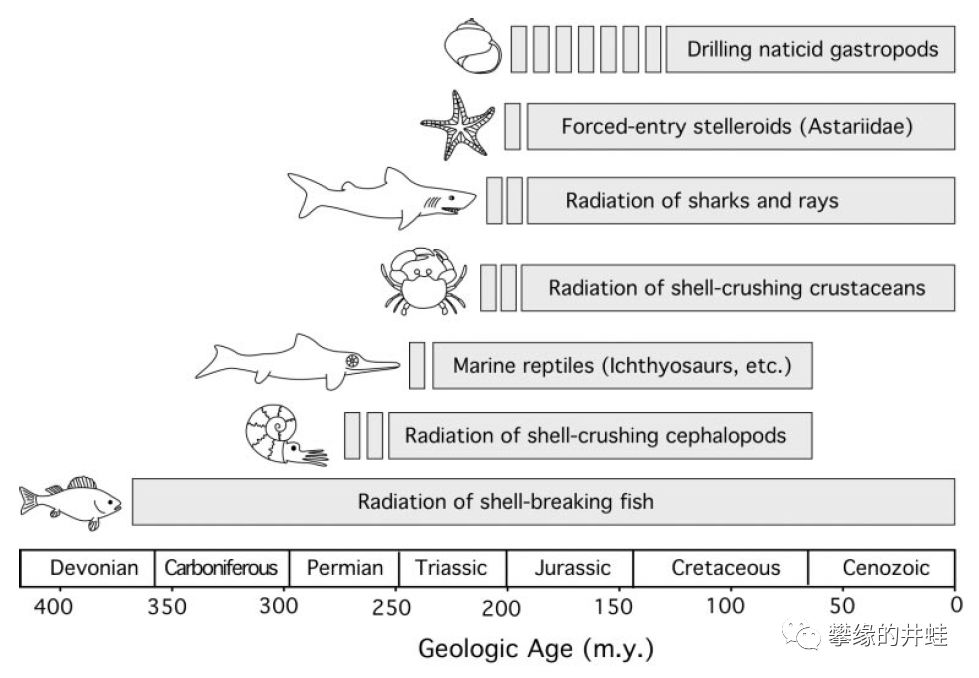
各种以双壳类(包括腕足动物)为食的动物类群的出现时间。三叠纪之后,与双壳类的繁盛同步,食贝动物的种类明显增多。图片来源自[10]。


直到今天,双壳类仍然是许多水生蜥形纲和合弓纲的主要食物。图片来源见水印。
地球名片
生物分类:动物界-软体动物门-双壳纲-蚌目
存在时间:石炭纪早期至现代
现存种类:约900种
化石种类:不明
生活环境:淡水,少数海洋
代表特征:除三角蛤科外大多淡水产,常具拟主齿或铰合齿退化。
代表群类:河蚌,珍珠蚌,三角蛤
参考文献:
[1] P.Yu. Parkhaev, Origin and the Early Evolution of the Phylum Mollusca.Paleontological Journal, 2017, Vol. 51, No. 6, pp. 663–686
[2] YutaShiino, Passive feeding in spiriferide brachiopods: An experimentalapproach using models of Devonian Paraspirifer and Cyrtospirifer. Lethaia43(2):223-231, August 2009, DOI: 10.1111/j.1502-3931.2009.00185.x
[3] SHIINO, Y. & KUWAZURU, O.,2011:07:29. Comparative experimental and simulation study on passive feedingflow generation in Cyrtospirifer. Memoirs of the Association of AustralasianPalaeontologists 41, 1-8. ISSN 0810-8889
[4] V. V. Silantiev, S. Chandra, M. N.Urazaeva, Systematics of Nonmarine Bivalve Mollusks from the Indian GondwanaCoal Measures (Damuda Group, Permian, India). Paleontological Journal, 2015,Vol. 49, No. 12, pp. 1235–1274
[5] Marcello GuimarãesSimões, Jacqueline Peixoto Neves, Luiz Eduardo Anelli, et al., Permian bivalvesof the Taciba Formation, Itararé Group, Paraná Basin, and theirbiostratigraphic significance. Geol. USP, Sér. cient., São Paulo, v. 12, n. 1,p. 7-82, DOI: 10.5327/Z1519-874X2012000100006
[6] V.V. Silantiev, Permian Nonmarine Bivalve Zonation of the East European Platform.Stratigraphy and Geological Correlation, 2014, Vol. 22, No. 1, pp. 1–27
[7] CHRISTOPHER A. McROBERTS, NORMAN D.NEWELL, Marine Myalinidae (Bivalvia: Pterioida) from the Permian of West Texas.American Museum Novitates, January 2005, Number 3469
[8] Ana Márquez-Aliaga, Antonio PabloJiménez-Jiménez, Antonio G. Checa, et al., Early oysters and their supposedPermian ancestors. Palaeogeography, Palaeoclimatology, Palaeoecology, 229(2005), 127–136, doi:10.1016/j.palaeo.2005.06.034
[9] Chenyi Tu, Zhong-Qiang Chen, David A. T . Harper, Permian–Triassic evolution of the Bivalvia:Extinction-recovery patterns linked to ecologic and taxonomic selectivity. Palaeogeography,Palaeoclimatology, Palaeoecology, Volume 459,October 2016, Pages 53-62, https://doi.org/10.1016/j.palaeo.2016.06.042
[10] Christopher A. McRoberts, Triassicbivalves and the initial marine Mesozoic revolution: A role for predators?Geology; April 2001; v. 29; no. 4; p. 359–362
感兴趣的话,不妨关注攀缘的井蛙,每天琢磨点新东西: