

细菌生物膜的特征及抗细菌生物膜策略
Bacterial biofilms: characteristics and combat strategies

来源

药学学报 Acta Pharmaceutica Sinica 2018, 53
作者
陈小楠, 申元娜, 李彭宇, 邹祎晴, 胡海燕
中山大学药学院
摘要
80%细菌感染与细菌生物膜 (简称菌膜) 形成有关, 相比于游离菌, 菌膜对抗生素的耐受性提高10~1 000 倍, 是造成目前细菌耐药的主要原因。了解菌膜的相关特征和耐药机制, 有助于更好地解决因菌膜导致的细菌感染及耐药性问题。
本文从菌膜的结构着手, 介绍了菌膜的结构、主要形成方式和菌膜的相关耐药机制, 在此基础上归纳了近年来发现的具有抗菌膜活性及与抗生素有协同作用的化合物, 并总结了新出现抗菌膜纳米递药系统, 以及分子印迹聚合物等抗菌膜新策略。
正 文
世界卫生组织(WHO) 称, 抗生素由于广泛、长期和不合理应用, 其耐药性已成为对全球健康和安全最严重的威胁之一, 使国际公共卫生面临前所未有的挑战。
细菌耐药性系指细菌对于抗菌药物作用的耐受性。耐药性一旦产生, 药物的化疗作用就明显下降。据统计, 80%细菌感染与细菌生物膜 (简称菌膜) 形成相关, 菌膜中的细菌形态和生理作用均与游离菌不同, 对抗生素的耐受性可以提高10~1 000 倍, 且对宿主免疫防御的抗性很强, 是造成细菌耐药性的主要原因[1]。
菌膜指细菌嵌入自身分泌的胞外聚合物(extracellular polymeric substances, EPS) 内形成的含有多细胞的三维结构群体。
EPS 包括多糖、蛋白质、核酸和脂类物质等, 是构成细菌生长的直接环境。为了生存, 细菌可以通过调节菌膜的结构以适应环境的变
化, 当菌膜暴露于亚抑制浓度的抗菌药物或金属离子等不利环境时, 可以诱导编码EPS 组分的基因表达, 促发EPS形成增多, 改变菌膜的三维结构使其向有利于生存的方向发展[2]。
大多数菌膜中, 微生物仅占干重的10%, EPS 占比却高达90%以上。根据微生物种类、所经历的剪切力和营养物质的获得性等差异, 不同细菌形成的菌膜EPS 差异很大。
例如, 从污泥中菌膜提取的EPS中, 蛋白质占43.5%, 腐殖酸类物质占11.5%, 核酸占10.9%, 多糖占9.9%, 脂质占1.8%[3]; 而从恶臭假单胞菌菌膜提取的EPS 中含蛋白质6%~12%、核酸13%~28%、多糖12%~44%及糖醛酸14%~58%[4]。
多糖和蛋白质是EPS 的主要成分, 常以接有凝集素的荧光染料标记多糖或接有特异性序列的染料标记蛋白质来指示EPS 的结构及含量。
多糖和蛋白质对于菌膜是不可或缺的, 多糖本身可以缔合钙镁等二价阳离子增强形成菌膜时的结合力, 而蛋白质能进一步稳定菌膜中多糖基质网络, 增强菌膜内的细菌与EPS 的连接[5]; 此外, 蛋白质还可以作为胞外酶参与降解EPS 促使细菌从菌膜分离, 并促进形成新的菌膜[6]。
胞外DNA (extracellular DNA, eDNA) 可作为细胞间连接分子、黏附素等存在于EPS 中, 在菌膜饥饿时可以作为营养源[7]。脂质在EPS 中所占比例很低, 但却对菌膜的黏附能力有重要影响, 并赋予了EPS 重要的属性——疏水性[3]。
菌膜除了内部细菌和EPS, 还存在一些信号分子充当细菌间通讯联络的信使。
当细菌密度增加时,细菌会加速自诱导分子的合成, 在达到一定的浓度阈值后, 通过调控细菌相关的基因表达来适应环境变化, 称为群体感应(quorum sensing, QS), 如调节菌膜的形成与分散[8]。
环二鸟苷酸(cyclic diguanylate, c-di-GMP) 是广泛存在于细菌中的第二信使, 调节细菌在游离状态与菌膜状态间进行转变。研究表明, 高浓度的c-di-GMP 能增加细菌的表面黏附与聚集, 以及加速EPS的分泌促进菌膜形成[9]。
QS 和c-di-GMP 调控菌膜的诸多生理功能, 然而两者重要性很少被同时提及。
菌膜一经形成即成为细菌的庇护所, 显著增强细菌对抗菌药物的抗性和对宿主免疫系统的逃避能力, 引发顽固性和复发性的感染。由于其组成结构和耐药机制的复杂性, 用传统的抗生素治疗很难将其彻底清除及杀灭, 采用增大抗生素用量的方法不仅对菌膜无效, 还会导致无法预期的毒副作用。
全面系统地认识菌膜, 有助于解决因菌膜引起的细菌感染性疾病及其耐药性问题。本文从剖析菌膜的结构着手,通过阐述其形成过程并分析其耐药机制, 综述了目前对菌膜的清除策略。
1
菌膜的形成
只要条件适宜, 任何细菌都可以形成菌膜。如图1 所示, 大部分菌膜黏附于活体组织或固体表面形成,称为经典附着型菌膜; 少部分细菌在没有底物的情况下也可以形成菌膜, 称为Ⅱ型非附着型菌膜[10]。
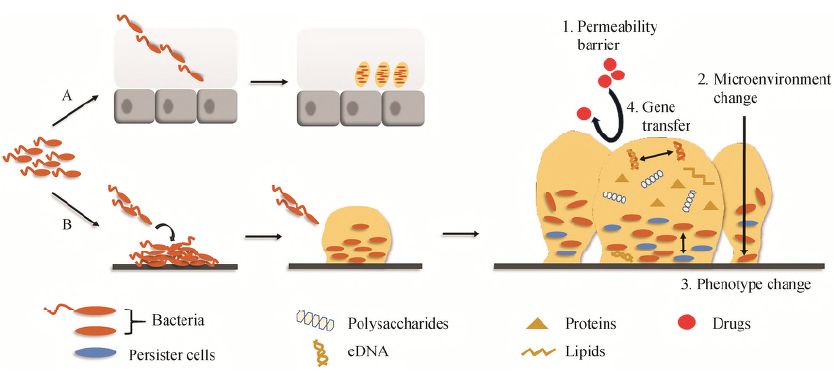
Figure 1 The formation, structure and resistance mechanisms of biofilm. A: A few bacteria can develop biofilm without substrate. Bacteria use flagella to actively penetrate into the mucus, and secrete extracellular polymeric substances (EPS) to wrap themselves, which are closely connected and enmeshed within the mucus. B: most biofilm need to adhere to living tissues or solid surfaces. The bacteria adhere to the surface through the flagella and cilia, and secrete EPS to gradually develop the mature biofilm. At this time, bacteria show obvious resistance to antibiotics. The mechanisms of drug resistance can be roughly divided into permeability barrier, microenvironment change, phenotypic change and gene transfer
经典附着型菌膜形成的主要步骤: 当游离菌感受到来自外界环境的生存压力后, 会通过鞭毛、纤毛等动力结构黏附到表面。此时, 单个附着的细菌仅由少量的EPS 包裹[11]; 随着细菌的增殖, 细菌分泌的QS 信号分子浓度达到一定阈值, 相关基因的表达开始发生变化, 并分泌大量EPS, 填充菌膜中细菌之间的空间, 为菌膜提供机械稳定性, 并逐渐形成蘑菇状或堆状的成熟菌膜, 此时细菌对抗生素表现出明显的耐药性[10]。
然而, 由于EPS 对物质交换的阻碍, 导致菌膜内部营养物质的缺乏及有毒代谢产物的堆积,细菌对该环境产生不适应, 这时菌膜会分泌胞外水解酶水解EPS, 破坏菌膜, 使细菌分散, 最终转化为游离状态[12]。
这些从菌膜脱离出来的游离菌在时机成熟时又可以重新黏附定植于其他位置, 发展成新的菌膜。
由于细菌存在游离菌—菌膜—游离菌—菌膜之间的不断循环转化, 最终引发持续性感染。
铜绿假单胞菌可嵌入厚厚的气道黏液内形成菌膜, 是经典的Ⅱ型非附着型菌膜。
囊性肺纤维化表现为外分泌腺的功能异常, 以及纤毛清除能力的下降,这导致管腔内黏液层逐渐变厚并形成缺氧梯度[13]。因此, 铜绿假单胞菌利用鞭毛主动渗透进入黏液层缺氧区, 或者借助黏液运动被动地包裹在黏液层内[14]。
在这种非附着状态下, 铜绿假单胞菌从有氧代谢转化为无氧代谢, 分泌EPS包裹自身, 嵌合在黏液层内部彼此紧密相连[15]。此时, 铜绿假单胞菌失去极端单鞭毛使得无法引发白细胞介素-8 的产生及嗜中性粒细胞的募集, 因此不被免疫系统识别而无法被清除[16]。
2
菌膜的耐药机制
菌膜对抗菌药物的耐药性通常是游离菌的10~1 000 倍, 是抗生素治疗细菌感染失败的主要原因。
菌膜的耐药机制复杂, 且不同机制之间存在协同作用, 主要有:
① 渗透屏障。
首先, 细菌分泌的EPS 形成致密并富集负电荷的天然屏障, 阻碍抗菌药物尤其是带正电荷的抗生素向菌膜内部渗透, 从而降低了菌膜内部抗菌药物浓度; 其次, 菌膜中的灭活酶可水解抗生素, 使其在渗透之前即在表层失活; 此外,混合菌膜结构致密, 使抗菌药物渗透难度远大于单一菌膜, 如白念珠菌与金黄葡萄球菌混合感染形成的菌膜耐药性显著增强[17]。
② 微环境改变。
菌膜中营养物质含量和氧气浓度等均自外向内逐步递减。营养缺乏使菌膜深层的细菌代谢速率减慢甚至进入休眠状态, 使其对繁殖期杀菌剂的抗性增强, 而菌膜内部缺氧亦会降低抗生素本身的杀菌能力[18]。此外, Liu等[19]研究发现, 菌膜具有震荡生长 (周期性生长) 的特点, 即外部细菌通过周期性的生长停止, 促使内部细菌获得营养从而保持其活性, 解决菌膜内外部的生长对营养物质需求的矛盾。最近, Martinez-Corral等[20]发现这种震荡生长的频率从小振幅开始, 随着细菌增殖而缓慢增长, 这允许菌膜能快速地应对外界条件的变化, 无疑增加了抗菌膜的难度。
③ 表型改变。
细菌为了适应环境, 通过特定基因表达改变致使其生物学行为发生改变。Whiteley团队[21]发现, 铜绿假单胞菌菌膜与游离菌比较有73 种基因表达呈现了差异, 尤其是普遍应激反应的相关基因及外排泵基因的表达量均高于游离菌, 并激活一系列基因表达抵御环境压力。另外, 菌膜中还存在一些因表型异化而产生能耐受多种抗生素的持留菌, 菌膜内部低氧和营养缺乏是促使持留菌形成的主要原因[22]。据报道[23]菌膜内部持留菌数量是游离菌的100~10 000倍, 是菌膜耐药的重要因素。
④ 基因平行转移。
菌膜内细菌通过质粒的接合完成基因的水平转移和吸收, 与游离菌相比, 菌膜中的质粒接合效率高达700倍[24], 其中, EPS 中的eDNA 可以作为水平基因转移的DNA 来源[25]。Lim 等[26]通过转录分析发现, 与质粒复制、基因交换等相关基因的表达量在菌膜形成初期会被上调, 如编码转座酶、整合酶和重组酶等基因的表达水平明显高于游离菌。
3
抗菌膜策略
在菌膜被认知之前, 抗菌策略主要聚焦于防止细菌定植。随着菌膜研究的深入, 以及菌膜对疾病发生发展作用的进一步阐明, 基于菌膜在结构及生活方式与游离菌显著的不同, 提出针对菌膜的治疗策略。
本文将能清除菌膜的代表性化合物、佐剂以及近年来方兴未艾的药物新型递药系统在抗菌膜中的应用进行综述。
3.1 清除菌膜的化合物及佐剂
EPS是菌膜的重要组成部分, 鉴于EPS对菌膜耐药性的重要贡献, 作用于EPS 和菌膜内部细菌均不失为清除菌膜的有效策略。
3.1.1 作用于 EPS
EPS 致密的结构构成了抗菌药物渗透的生理屏障, 加上抗菌药物被EPS捕捉并滞留, 极大地降低了抗菌药物对细菌的杀伤作用。因此, 作用于EPS 将成为清除菌膜的有效策略。
针对EPS 主要成分蛋白质、多糖、脂质和eDNA等, 可根据各种成分性质的不同对EPS 进行清除或破坏, 不同酶对不同菌膜的清除效果差异很大。
Pel 和Psl 是铜绿假单胞菌EPS 中的关键组分, Baker 等[27]研究发现, Pel和Psl生物合成操作子中分别编码的糖苷水解酶PelA(h) 和PslG(h) 在适宜的浓度下能迅速破坏成熟菌膜, 因而增强多黏菌素E 的抗菌作用。
而分散素B (dispersin B) 能分解鲍氏不动杆菌等多种菌膜EPS 中的重要成分1,6-乙酰-d-氨基葡萄糖聚合物。
Chen 等[28]研究表明, 将分散素B 与银结合肽融合后, 菌膜的清除率提高了20 倍。
除上述几种针对EPS 中多糖的酶外, 蛋白酶和脱氧核糖核酸酶也同样能破坏菌膜。如木瓜蛋白酶和胰蛋白酶能有效消化口腔放线菌菌膜中的主要纤维蛋白FimP 和FimA,并降低了混合菌膜的体外重建率[29]。
此外, 表面活性剂的抗菌膜研究近年来受到了广泛关注。由于表面活性剂带有电荷并具有两亲性,可通过与菌膜发生相互作用破坏EPS 的结构与功能。
如柠檬酸两性离子表面活性剂, 能螯合EPS 中的钙离子桥, 使EPS 结构的稳定性降低, 从而清除菌膜[30]。当十二烷基硫酸钠达到临界胶束浓度后, 可影响EPS 中蛋白的折叠甚至引起变性, 从而使菌膜瓦解。
此外, 以鼠李糖脂为代表的生物表面活性剂多有抑菌作用, 能削弱细菌的黏附, 抑制菌膜的形成。鼠李糖脂还可以调节菌膜的信号通路, 引起菌膜解体[31]。
除了酶和表面活性剂外, 最近有研究使用人工模拟细胞清除菌膜。Martin 团队[32]把识别铜绿假单胞菌信号分子的传感器基因、内酯酶基因和糖苷水解酶基因共同转染进人工模拟细胞进行表达, 实验发现转染后的人工模拟细胞可以很好地分散及清除铜绿假单胞菌菌膜, 并降低毒力因子的表达。
将人工模拟细胞与妥布霉素联用可以增强菌膜对妥布霉素的敏感性。而他们在后续的研究中还将抗菌肽基因也转染至人工模拟细胞中, 直接杀死菌膜内的细菌[33]。
此外, 表面活性剂的抗菌膜研究近年来受到了广泛关注。由于表面活性剂带有电荷并具有两亲性,
可通过与菌膜发生相互作用破坏EPS 的结构与功能。如柠檬酸两性离子表面活性剂, 能螯合EPS 中的钙离子桥, 使EPS 结构的稳定性降低, 从而清除菌膜[30]。
当十二烷基硫酸钠达到临界胶束浓度后, 可影响EPS 中蛋白的折叠甚至引起变性, 从而使菌膜瓦解。
此外, 以鼠李糖脂为代表的生物表面活性剂多有抑菌作用, 能削弱细菌的黏附, 抑制菌膜的形成。鼠李糖脂还可以调节菌膜的信号通路, 引起菌膜解体[31]。
除了酶和表面活性剂外, 最近有研究使用人工模拟细胞清除菌膜。Martin 团队[32]把识别铜绿假单胞菌信号分子的传感器基因、内酯酶基因和糖苷水解酶基因共同转染进人工模拟细胞进行表达, 实验发现转染后的人工模拟细胞可以很好地分散及清除铜绿假单胞菌菌膜, 并降低毒力因子的表达。
将人工模拟细胞与妥布霉素联用可以增强菌膜对妥布霉素的敏感性。而他们在后续的研究中还将抗菌肽基因也转染至人工模拟细胞中, 直接杀死菌膜内的细菌[33]。
3.1.2 作用于菌膜内细菌
3.1.2.1 群体感应抑制剂
阻止自诱导分子积累、促进其降解或影响其与受体的识别和结合, 可以抑制QS,此称为群体淬灭。开发QS 抑制剂(quorum sensing inhibitors, QSI) 有望有效治疗菌膜相关感染。
以酰基高丝氨酸内酯(acyl-homoserine lactones, AHLs) 为例, 迄今为止, 许多AHL-内脂酶和AHL-酰基转移酶被发现可以降解AHL使其失活而阻断QS[34], 最近研究表明, 由于与AHL-酰基酶具有一定的同源性, 在制药工业应用广泛的青霉素酰化酶也显示有AHL 降解活性[35]。
此外, Ma 等[36]从南海的珊瑚群落中分离出约200个细菌群落, 进行筛选后发现约15%分离菌株对指示菌株 (紫色杆菌或铜绿假单胞菌) 有抗QS活性。
在此基础上, QSI 与抗生素联合使用还可以提高菌膜内细菌对抗生素的敏感性。Busetti 等[37]发现海洋提取物KS8 表现出强烈的QSI 活性, 其上清液能抑制约60%的铜绿假单胞菌菌膜形成, 对成熟菌膜的清除率达30%。
KS8 与妥布霉素联用可使妥布霉素的疗效提高10 倍。Vermote 等[38]则发现金缕梅单宁类似物能显著增加金黄色葡萄球菌对抗生素的敏感性。
该物质与万古霉素、头孢唑林、达托霉素和妥布霉素等抗生素联用时, 清除菌膜的能力可提高至少20 倍。
3.1.2.2 外排泵抑制剂
菌膜产生耐药性的另一个原因是外排泵的过度表达。针对细菌外排泵的药物设计将成为抗菌治疗的一个重要策略。
通过生物信息学分析, Sirijant 等[39]发现, 在伯克霍尔德菌中, 头孢他啶、多西环素和亚胺培南的最低菌膜抑制浓度和最低菌膜清除浓度是游离菌的2~1 024 倍。
当添加外排泵抑制剂苯丙氨酸−精氨酸-β-萘胺后, 头孢他啶和多西环素的最低菌膜清除浓度降低为原来的1/16 至1/2。
由此可见, 抑制外排泵可以使菌膜对头孢他啶和多西环素的耐药性降低。
此外, 使用以拓扑异构酶IA 为靶点的广谱抗生素PPEF 后引起了大肠杆菌K12 外排泵基因的表达上调, 而当同时使用外排泵抑制剂羰基氰化物3-氯苯基肼(carbonyl cyanidem-chlorophenylhydrazone, CCCP) 后, 产生了显著的协同抗菌膜作用[40]。
3.1.2.3 抗菌肽
抗菌肽(antimicrobial peptide, AMP)一般由20~60 个氨基酸残基组成, 是具有抗菌活性的一类碱性多肽物质。
天然的AMP 来源非常广泛,具有高效广谱的杀菌特性, 通过重组或改造可使天然AMP 抗菌能力得到增强。
LL-37 是Cathelicidin 家族中唯一存在于人体的AMP, Overhage 等[41]发现LL-37 能有效抑制铜绿假单胞菌菌膜的形成, 对预先形成的菌膜也有一定的清除作用。
而Mishra 等[42]基于LL-37 的三维结构设计了17BIPHE2 AMP, 研究发现17BIPHE2 和抗生素联用对成熟铜绿假单胞菌菌膜能达到较高的清除率, 效果优于天然LL-37。
此外, de la Fuente-Nunez等[43]还发现了一种有效的抗菌肽1018 AMP, 在低于最低抑菌浓度下, 能完全抑制耐甲氧西林金黄葡萄球菌、绿脓杆菌、大肠杆菌和鲍曼不动杆菌等菌膜的形成。并且, 低浓度1018AMP 可致菌膜解体, 高剂量1018 AMP 可致菌膜内
细菌死亡。
3.1.2.4 代谢促进剂
菌膜内处于休眠状态的持留菌数量是环境中的10~10 000 倍, 使菌膜能在高浓度抗生素中存活。若将持留菌唤醒, 使其恢复为代谢活跃的状态, 则可以提高菌膜对抗生素的敏感性[44]。
Allison 等[45]发现, 氨基糖苷类抗生素与特定的中间代谢物 (葡萄糖、甘露醇、果糖、丙酮酸等) 联合应用可以杀死大肠杆菌和金黄葡萄球菌菌膜, 然而这种用中间代谢物来刺激代谢的方法并不能迅速使持留菌恢复到细胞壁合成和DNA合成活跃的生长状态。
研究进一步发现, 由于这些特定的中间代谢物诱导了质子动力, 促进了氨基糖苷类抗生素的摄取最终才致使细菌的死亡。
因此, 将诱导质子动力增强的代谢产物作为佐剂与氨基糖苷类抗生素联用进行菌膜治疗理论上是可行的。
此外, Marques 等[46]证实顺2-癸烯酸 (cis-2-decenoic acid, cis-DA) 这种脂肪酸信号分子可以使铜绿假单胞菌和大肠杆菌的持留菌从休眠状态转化为代谢活跃状态。
将cis-DA 分别与妥布霉素、四环素联合使用, 可以显著降低细菌的存活率。
3.2 药物递送系统
3.2.1 纳米粒
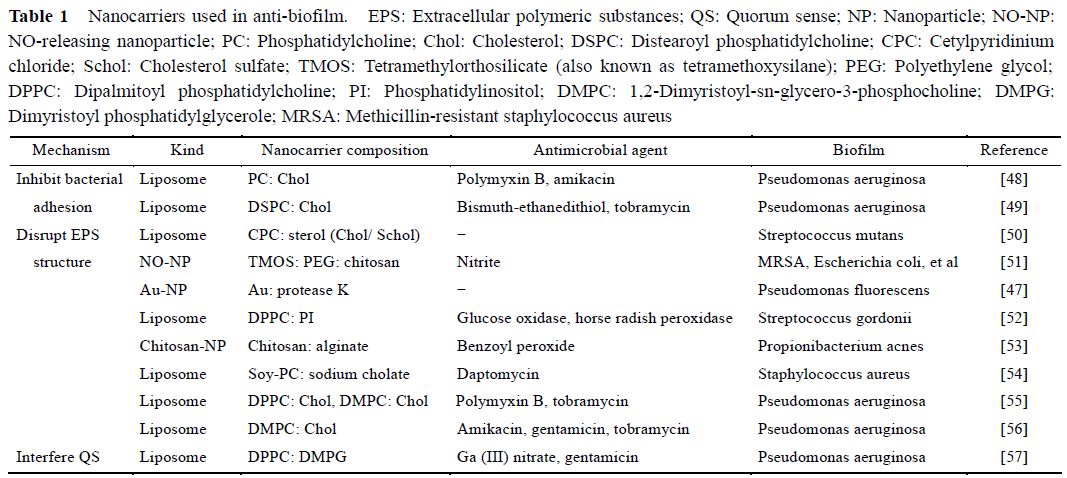
最近十几年来, 采用纳米递药系统来抑制菌膜形成及清除成熟菌膜受到越来越多的关注, 同时被认为是抗菌膜感染的有效策略。根据材料来源的不同,将抗菌膜的纳米载体分为无机纳米载体及脂质纳米载体两大类。
表1[47−57]总结了采用不同作用机制递送系统抗菌膜的例子。
3.2.1.1 无机纳米载体
近几年使用无机纳米粒对抗菌膜的研究相对较多。金属纳米粒是抗菌膜研究的热点, 除具有较强的抗菌活性外, 还显示较强的抗菌膜效果。
金属粒子如银、金、锌、铜、钛和铋等主要是通过损伤细菌中的DNA、蛋白质等大分子, 破坏细菌的正常结构达到抗菌目的。
金属纳米粒表面可以通过修饰配体来增强抗菌膜能力和靶向性。如Habimana 等[47]在金纳米粒上修饰蛋白酶K, 蛋白酶K首先破坏EPS结构, 菌膜内细菌暴露, 金纳米粒发挥杀菌作用。结果表明, 使用单独的蛋白酶K、金纳米粒及蛋白酶K 修饰的金纳米粒处理荧光假单胞菌菌膜, 对菌膜的清除能力分别达到40%、77%及 74%清除率, 虽然蛋白酶K 修饰的金纳米粒清除菌膜并非最优, 然而经其处理后的菌膜内部细菌存活率最低。
此外, 在金属纳米粒上连接靶向性的基团, 可以实现主动靶向的能力。
Bhargava 等[58]在银纳米粒表面连接L-岩藻糖, 可与铜绿假单胞菌的凝集素结合实现主动靶向, 然后利用银优良的抗菌特性来杀菌。研究表明, 载有L-岩藻糖的银纳米粒能有效抑制铜绿假单胞菌菌膜的形成,并杀死菌膜内部82.6%细菌。
除金属纳米粒外, 目前以二氧化硅、氧化石墨烯等无机材料为载体的制剂在抗菌膜领域也有研究。
Duan 等[59]将β-内酰胺酶抑制剂舒巴坦载入介孔二氧化硅纳米粒中, 外层包被具有pH 敏感作用及门控作用的三价铁离子与β-内酰胺类抗生素羧苄青霉素螯合物。
当其到达pH 较低的菌膜区域时, 羧苄青霉素与三价铁离子解离, 促使内层的舒巴坦释放。这种核−壳结构在生理条件下是稳定, 能在菌膜定植部位释放抗生素和抑制剂, 从而更好地治疗因菌膜引起的感染。
而氧化石墨烯比表面积高、易于改性并且高度亲水性, 是抗菌药物的理想纳米载体。
Zhao 等[60]将氧化石墨烯与纳米银离子组合起来协同抗菌, 并在表面修饰水溶性高分子聚合物聚乙烯亚胺使其兼具稳定性好且长效的优点。研究表明, 构建的复合物贮藏1 周后在相同浓度下时仍能杀灭99%以上的微生物 (包括耐药细菌及真菌), 且抑制90%菌膜形成。
在此基础上, 还可将无机纳米粒与物理手段(如光热疗法) 联用增强抗菌膜的作用。
其原理是某些材料在光照刺激下可产生大量热量以破坏菌膜并杀死菌膜内的细菌。金在光照下可通过等离子共振效应产生大量热量, 越来越多的研究者将金纳米粒联合光热疗法用来清除菌膜。Hu 等[61]通过在金纳米粒表面修饰11-巯基十一酸(HS-C10-COOH) 和(10-巯基癸基) 三甲基溴化铵(HS-C10-N4) 使其具有pH 敏感特性。
此纳米粒在菌膜感染的低pH 环境下表面基团解离而带正电荷, 与带负电菌膜发生强相互作用。
并在光照的刺激下产生热量杀死并清除菌膜内的细菌。此外, 外加磁场也可以使纳米粒靶向至菌膜感染区域。
直径小于30 nm 的超顺磁性纳米粒利用其粒径小及磁响应特点, 可以渗透进菌膜, 解决纳米粒在EPS 中难渗透及菌膜清除率低的问题。
Taylor等[62]和Durmus等[63]在超顺磁性纳米粒表面耦合3种不同的金属离子 (铁、银和锌), 与抗生素和金属盐(抑制游离菌但促进菌膜生长) 比较, 耦合了金属离子的超顺磁性纳米粒对游离菌及菌膜均有抑制作用。
且在外加磁场下, 耦合银离子的超顺磁性纳米粒能更好地清除菌膜。
3.2.1.2 脂质纳米载体
已有越来越多使用脂质纳米载体抗菌膜的报道。一方面, 这些载体有较好的生物相容性、材料表面可修饰等优点; 另一方面, 脂质纳米载体可与菌膜互相融合实现菌膜靶向, 提高抗菌药物对菌膜的敏感性, 提高治疗效率降低不良反应[64]。
脂质纳米载体以脂质体最常见。目前主要分为两类,
一类是融合型脂质体, 其与菌膜外部融合并将药物释放进菌膜内部杀菌。研究表明, 包裹有氨基糖苷类抗生素的二棕榈酰磷脂酰胆碱或二硬脂酰基磷脂酰胆碱与胆固醇组成的脂质体在铜绿假单胞菌菌膜与洋葱伯克霍尔德菌菌膜中具有更好的抗菌活性,并通过透射电镜、流式细胞术及脂质混合实验等证明了脂质体与细菌膜的融合[65, 66]。
另一类是黏附性脂质体, 利用脂质体所带的正电荷与负电性的菌膜相互作用, 黏附在菌膜上持续释放药物杀死内部的细菌。
Alhajlan 等[67]以9 种铜绿假单胞菌菌株为研究对象,测试将克拉霉素分别包封在阴性 (带负电)、阳性 (带正电) 及中性 (不带电) 脂质体时的抗菌活性。研究发现, 阴性脂质体与阳性脂质体能完全清除菌膜, 但阳性脂质体有效浓度相对较低。这可能由于带有正电荷的脂质体与带有负电的菌膜静电吸引, 持续接触菌膜使脂质体更好地释放药物进入菌膜内部, 提高药物到达细菌部位的浓度。此外, 在脂质体上修饰靶向基团小麦胚芽凝集素, 可以选择性地与EPS 中的N-乙酰氨基葡萄糖等结合, 提高脂质体对菌膜的靶向性结合[68]。
目前, 许多公司正在开发用于治疗菌膜感染的脂质体制剂, 一些正在进行临床试验, 部分已经上市。
如相较于游离的妥布霉素, 以二肉豆蔻酰磷脂酰甘油和二棕榈酰磷脂酰胆碱为脂质载体的妥布霉素脂质体Fluidosomes 能显著降低铜绿假单胞菌菌膜的存活, 并且在给药后大部分停留在肺部, 提高了疗效,降低了毒副作用[69]。
Fluidosomes 目前由Axentis 制药研发, 暂未获得该品种临床试验的进一步信息。阿米卡星脂质体Arikace是另一种用于治疗由铜绿假单胞菌菌膜引起的肺部疾病的脂质体。
脂质体外壳保护带正电的阿米卡星不受患者痰液中负电荷成分的影响,当脂质体到达感染部位后, 会被分泌的鼠李糖脂裂解, 脂质体内的阿米卡星释放出来。
Arikace 在2015年被美国FDA 指定为孤儿药, 用于治疗由铜绿假单胞菌和非结核分枝杆菌引起的肺部疾病[70]。
脂质聚合物纳米粒由外层脂质和内核聚合物组成, 同时具备外层脂质的生物相容性、聚合物内核的稳定及药物持续长效释放等优点, 并且物理化学性质较脂质体稳定。
Cai 等[71]以磷脂和鼠李糖脂为脂质外层, 果胶硫酸酯聚合物和阿莫西林为亲水内核, 构建用于清除幽门螺杆菌菌膜的脂质聚合物纳米粒,其外层脂质破坏菌膜EPS 结构分散细菌, 脂质聚合物纳米粒外层解体后释放阿莫西林杀菌, 暴露出果胶硫酸酯聚合物遇水溶胀抑制细菌的再黏附。
研究表明, 载阿莫西林脂质聚合物纳米粒能杀死菌膜内80%以上的细菌。而Cheow 等[72]以磷脂为外壳, 聚乳酸-羟基乙酸共聚物为内核制备了菌膜触发释放的脂质聚合物纳米粒, 铜绿假单胞菌菌膜中由于存在的鼠李糖脂, 其可以破坏脂质聚合物纳米粒的外脂层,从而控制药物的释放。
3.2.2 表面分子印迹载体
分子印迹技术(molecular imprinting technique,MIT) 指以某一特定的目标分子为模板, 制备对该分子具有特异选择性聚合物的过程。随着MIT 的不断发展, 分子印迹聚合物(molecular imprinted polymers,
MIPs) 已被用于设计新型药物传递系统。
由于结合位点可随着外界环境的变化而改变, 使设计智能MIPs通过响应特定生物分子变化来释放药物成为可能[73]。目前有两个团队以分子印迹达到抗菌膜的目的。
一是英国克兰菲尔德大学的Piletska 团队[74], 他们通过计算机模型以铜绿假单胞菌菌膜的自诱导分子3-oxo-C12-AHL 为模板进行MIP 的设计, 结晶紫染色定量检测菌膜含量, 发现设计的MIP 能抑制80%以上的铜绿假单胞菌菌膜的形成;
另一个团队是加拿大不列颠哥伦比亚大学Ma 等[75]同样以3-oxo-C12-AHL 为模板进行MIP 的合成, 并进一步优化了合成过程中功能单体的选择。
然而, 目前用于抗菌膜的都是空白MIPs (没有载药) , 并且只探讨了其抑制菌膜形成的能力, 没有探究其清除菌膜的能力。
复旦大学的潘俊教授课题组[76]基于MIT 构建了抗幽门螺杆菌生物黏附的纳米粒, 以幽门螺旋杆菌表面抗原片段NQA 和阿莫西林为双模板, 利用反相微乳液聚合法进行制备, 在体外成功实现对幽门螺旋杆菌的特异性结合, 证实菌膜特异分子和抗菌药物为双模板构建特异靶向菌膜的MIPs 是可行的。
MIPs 的主要优点是它们对印迹过程中使用的靶分子具有高选择性和亲和力。
与蛋白质和核酸等生物系统相比, 分子印迹聚合物具有更高的物理稳定性、强度、耐高温高压性和对酸、碱、金属离子和有机溶剂表现为惰性。此外, 它们的合成成本低, 储存寿命长, 即使在室温下也可以保持其特异识别能力达数年之久[77]。
4
结语
对细菌有直接杀伤作用的化合物 (如抗生素) 都有潜在的耐药危机。虽然DNA 测序、基因挖掘等技术为寻找新抗生素提供了新的思路和高效的路径[78],抗生素发现的速度和种类仍远远不能满足临床的需求。
因此, 针对菌膜特点设计的抗耐药菌策略, 如干扰菌膜QS或通过降解EPS直接破坏菌膜结构等方法为治疗菌膜引发的耐药性细菌感染提供了有效的手段。
考虑到菌膜破坏后会增加细菌扩散、传播及定植在新的感染部位重新形成菌膜的风险, 在采用降解EPS 策略时可联用抗菌肽、代谢促进剂等, 在破坏菌膜结构使游离菌充分暴露的基础上, 增强抗生素的敏感性, 并抑制菌膜再形成, 达到提高清除率降低复发率的目的。
在此基础上, 纳米制剂还可以实现对药物的控制释放, 进一步提高抗菌膜效果。
目前, 针对医疗设备表面修饰抗黏附药物或治疗皮肤表皮菌膜感染的研究已经相对成熟, 而由于体内环境的特殊性与复杂性, 针对体内菌膜感染的研究仍有限, 故迫切需要建立能准确模拟人体病理环境的体内外模型,评估抗菌膜递药系统的体内效果, 为后续新型递药系统的临床研究提供基础。
此外, 目前只有铜绿假单胞菌、葡萄球菌、肠球菌、链球菌和大肠杆菌等菌膜的研究报道, 但实际只要条件适宜, 任何细菌都可以形成菌膜。因此, 对其他菌膜的研究也应加强。