

当石炭纪的巨虫们凋零后,无脊椎动物中能够和脊椎动物刚正面,竞争高级掠食者地位的,就只剩下软体动物门头足纲(Cephalopoda)的一众触手怪了。

头足纲是无脊椎动物的巅峰也是异类。它们和其他软体近亲是如此不同,许多特征简直就像是为了抗衡(有颌类)脊椎动物专门设计的:强壮灵活的腕;复杂而灵敏的眼睛和其他感官;高效而节能喷射推进系统;角质喙和齿舌配合,可以轻易碾碎猎物的血肉和骨骼;还有功能强大的脑来指导复杂的行为甚至学习新技能。它们展示了高等动物的另一种演化可能。图片来源见水印。
研究者基本认同头足纲起源于某种寒武纪早期的,背着一片小贝壳的,外形像单板纲或者腹足纲的小型软体动物,也许就是太阳女神螺类。但没有人能从众多的小壳化石中把这种动物找出来——因为缺少软体结构的资料,谁也不知道挤在贝壳里面的动物到底更像蜗牛还是章鱼。
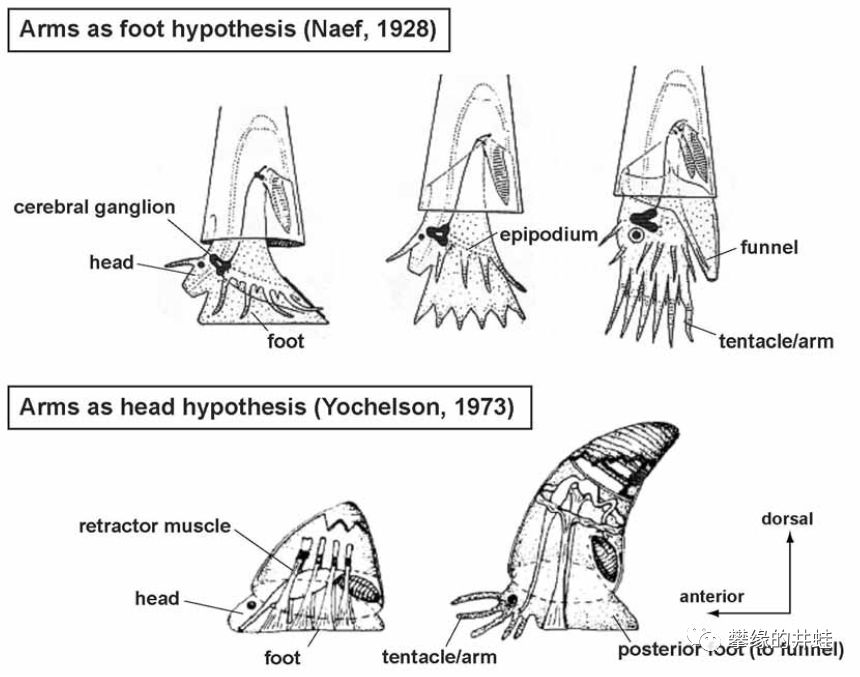
关于头足纲触手(腕)起源的两种解释。上:足源说,腕由腹足分裂而成。下:头源说,腕由头——准确地说是口周围的结构分裂强化而成。图片来源自[1]。
能够确认的最早的头足纲动物化石出现在寒武纪晚期,比如短棒角石Plectronoceras。它们的贝壳里已经形成了分隔的气室,体室和腹管,这些是头足类特有的结构。
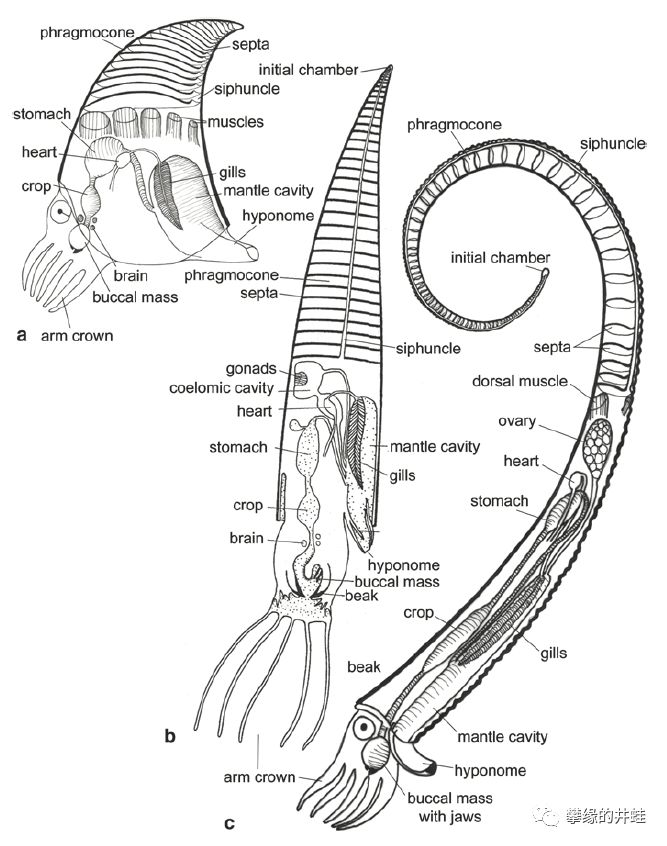
一些早期头足类的身体结构示意图。(a)是短棒角石,可以确认的最早的头足动物之一。(b)被称为“HASC(Hypothetical Ancestral Siphonopodean Cephalopod)”,井蛙译作“管足类头足动物的理论祖先”,它的后代包括鹦鹉螺,菊石和蛸形类。(c)是最早的菊石Metabactrites fuchsi。有壳头足类并不像其他软体动物那样,内脏团塞满整个贝壳。它们的贝壳被隔板(Septa)分隔成一层一层的,身体(包括鳃,消化道,生长系统等内脏)只呆在最下层体室(Chamber)里。体室之上的部分称为闭锥(Phragmocone),由多层气室(Chamber)组成,有一根腹管(Siphuncle)贯穿体室和气室,直通壳顶。因此发现了隔板,气室和腹管痕迹的贝壳,就可以确认是头足动物了。图片来源自[2]。
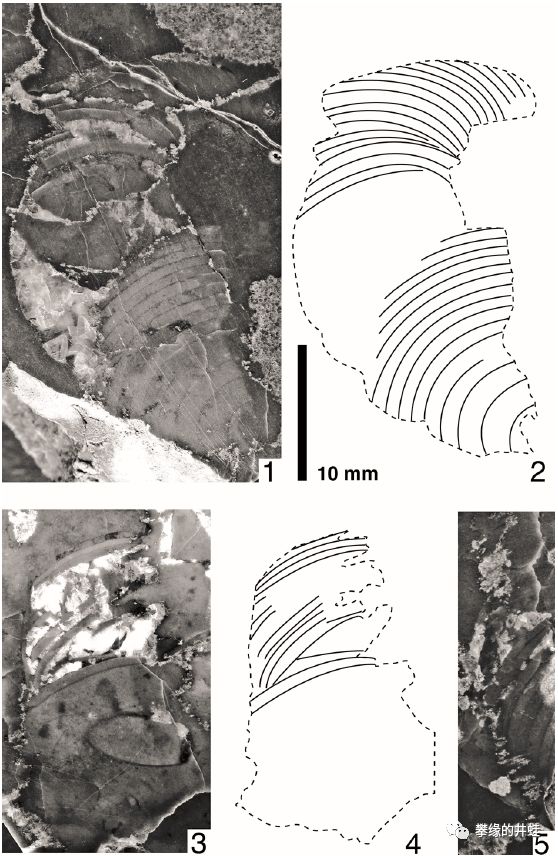
上图是纽约州发现的晚寒武世软体动物的贝壳,学名未定。上面有明显的隔板和气室痕迹,这是目前发现的最早的头足动物贝壳化石。图片来源自[3]。
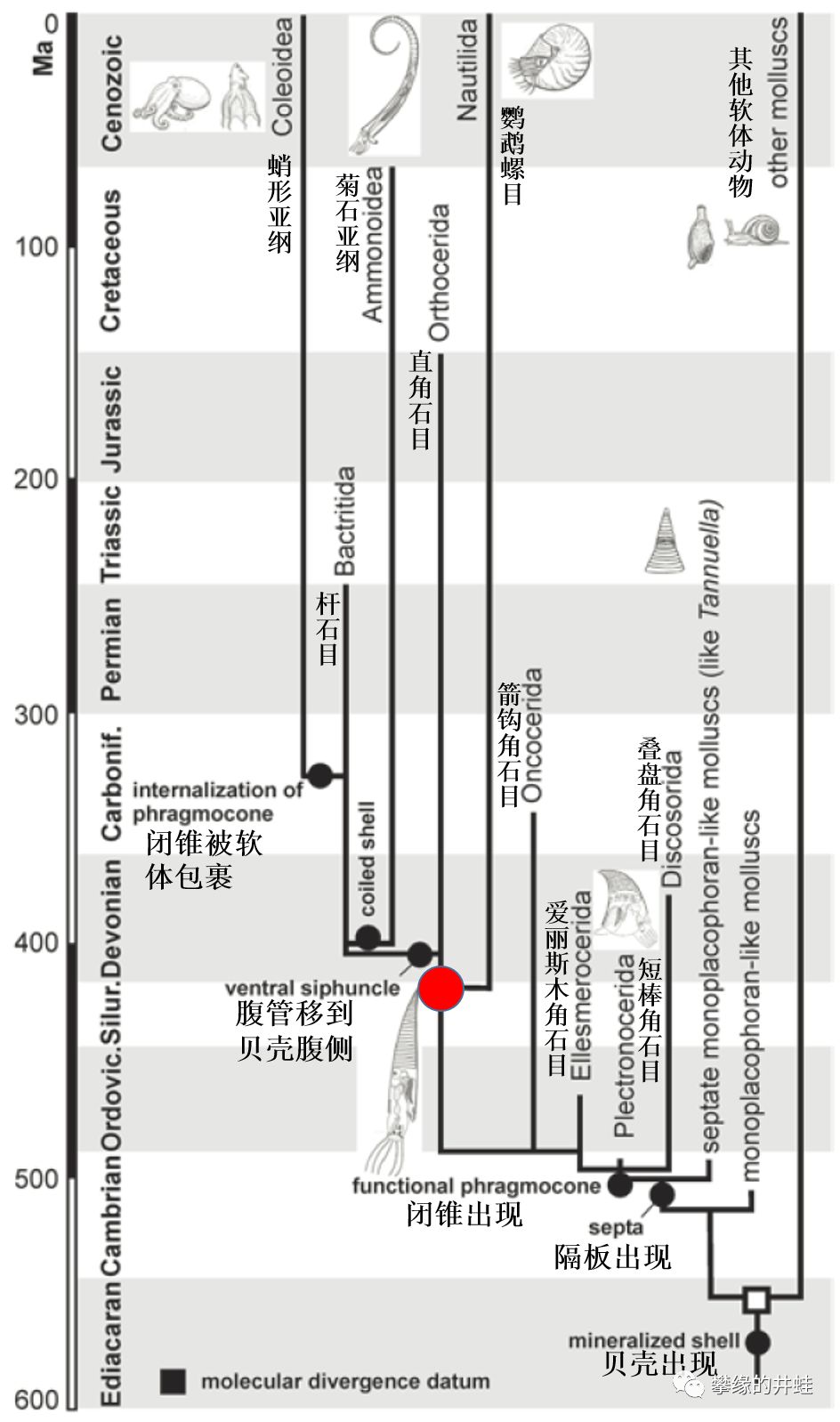
上面提到了一个很有意思的概念:HASC(Hypothetical Ancestral Siphonopodean Cephalopod,管足类头足动物的理论祖先)。它是研究者根据已知头足类的特征反推出的一个模型,是所有现存头足类(乌贼,章鱼和鹦鹉螺)最近的共同祖先。最接近这个模型的动物的演化地位和出现时间在上图中用红点标出。图片来源自[2]。
头足纲在奥陶纪飞速发展,内角石目的巨怪房角石Cameroceras一跃成为海洋的霸主。奥陶纪末的大灭绝事件并没有打断头足纲的繁荣,各种角石,鹦鹉螺和早期菊石在志留纪和泥盆纪继续兴旺发达。

在志留纪和泥盆纪,就算面对板足鲎和有颌鱼类的竞争,头足纲依然涌现出不少体型相当大的种类。上图是几种志留纪和泥盆纪直角石完整贝壳的大小(推测)。注意这只是贝壳,想象动物的实际尺寸,请在锥形的底部自行加上一个挥舞着触手的乌贼头。图片来源自[4]。

一些志留纪到泥盆纪的直角石类化石,产地:摩洛哥。它们巨大的锥形贝壳很难完整保存,这些碎片上有密集的隔板和气室,是靠近贝壳顶端的部分碎片。图片来源自[4],标尺长度1厘米。
直不隆捅的锥形和杆形外壳有许多缺点,比如尖细的末端很脆弱,容易破损。密闭的气室漏水,会妨碍头足动物调节浮力,影响运动,更让病原体和寄生物有机可乘。另外,头足动物喷水推进身体,细长的壳体控制和转向都十分困难。也许正因如此,直角石从石炭纪开始衰落,继之而起的鹦鹉螺和菊石分别演化出螺旋形的外壳,坚固程度和可操控性都大为提高。

一些泥盆纪中期鹦鹉螺化石。产地:德国Kluterthöhle岩洞。这些鹦鹉螺处在演化的早期,贝壳弯曲成各种形状,并没有完全盘曲成螺旋形。图片来源自[5]。

根据Kluterthöhle岩洞发现的化石复原的生态系统复原图,鹦鹉螺是主角。幸好它们的后代有几种生存到今天,人们才知道眼睛,触手和口盖大致的样子。图片来源自[6]。
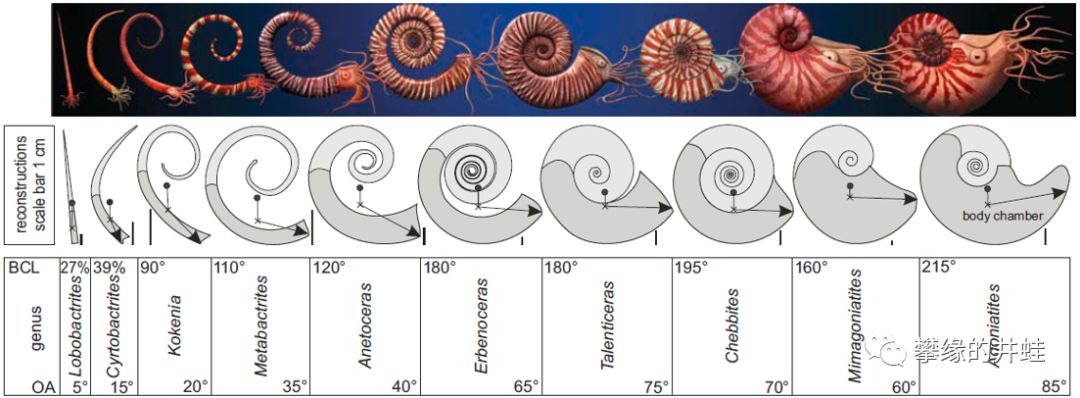
一些泥盆纪杆石和菊石的贝壳结构。它们和鹦鹉螺一样,留下了从尖锥形壳到螺旋形壳的各个演化阶段的化石。这也算是一种趋同演化吧。贝壳螺旋化的程度越高,就越坚固,游动速度和灵活性也越好。另一个演化趋势是体室所占的比例越来越大,为触手,眼睛,喙和脑提供了充足的发展空间。图片来源自[7]。
之前介绍过怎样根据缝合线区别鹦鹉螺和菊石(参见第一百三十四回克苏鲁之裔3:菊石之海)。本回如果再把一堆角石,鹦鹉螺和菊石的贝壳像流水账一样摊出来,大家也会觉得无聊吧?井蛙就挑一些有意思的化石来说说。
这是在美国蒙大拿州熊谷石灰岩(The BearGulch Limestone)发现的一块石炭纪菊石化石:

两个小小的贝壳并排平躺在一起,螺口相对。它们是正在进行生命大和谐时突然被掩埋的。这对小情侣就这样拥抱依偎着,在地层中沉睡了3亿年。图片来源自[8]。
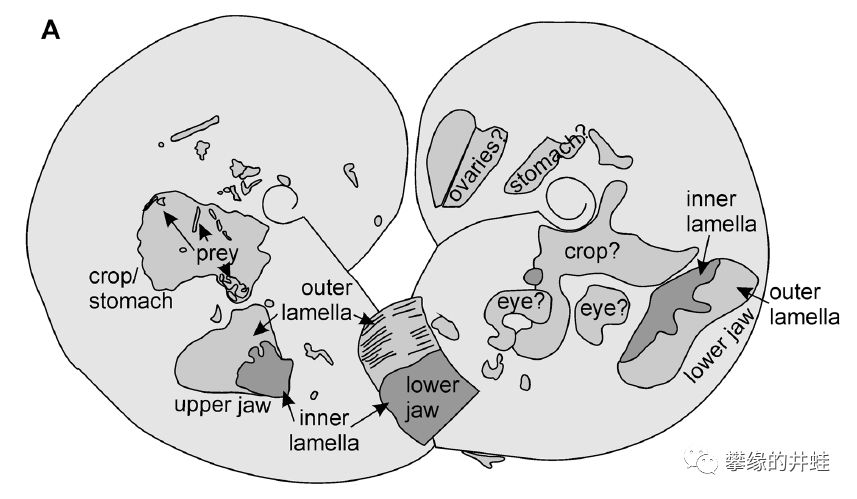
化石上有一些深色的斑块,可以识别出部分内部结构,包括喙,眼睛,消化系统等等。菊石的软体结构极其罕见,而且几乎全是这种模糊不清的印痕。图片来源自[8]。
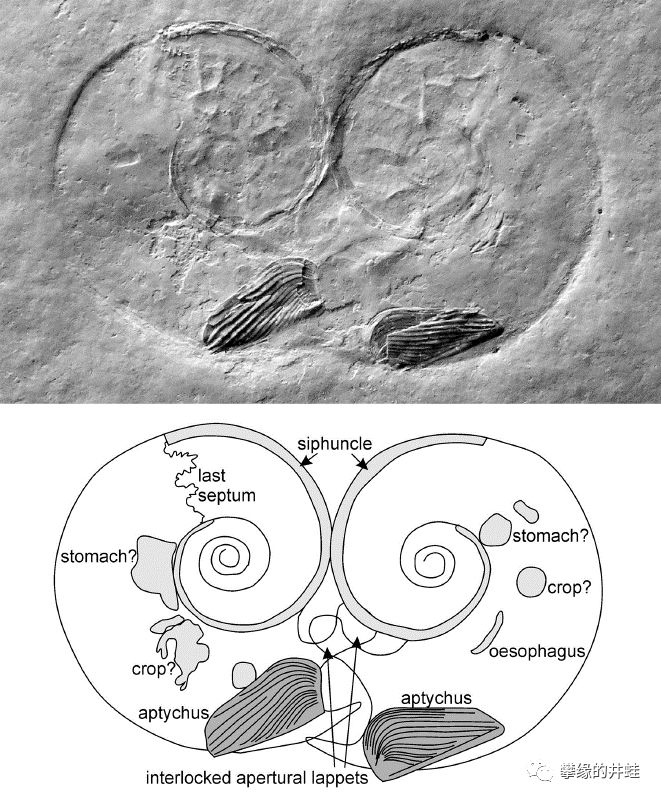
无独有偶,这块晚侏罗世的菊石化石也记录下了同样的瞬间,它们用自己的身体比了个心。它们比石炭纪的前辈多了口盖(Aptychus)的结构。发现地:德国Eichstätt。图片来自[8]。

现代鹦鹉螺也继承了这种啪啪啪的方式(好像多了一只?),几十条触手纠缠在一起,难解难分。图片来源见水印。
除了软体,贝壳表面的细微结构也会因风化溶蚀而丢失。幸好极少数化石上保留了一点颜色和纹饰的残迹,这些信息加上现代鹦鹉螺作参考,让研究者在复原古代头足动物时不至于完全乱猜。
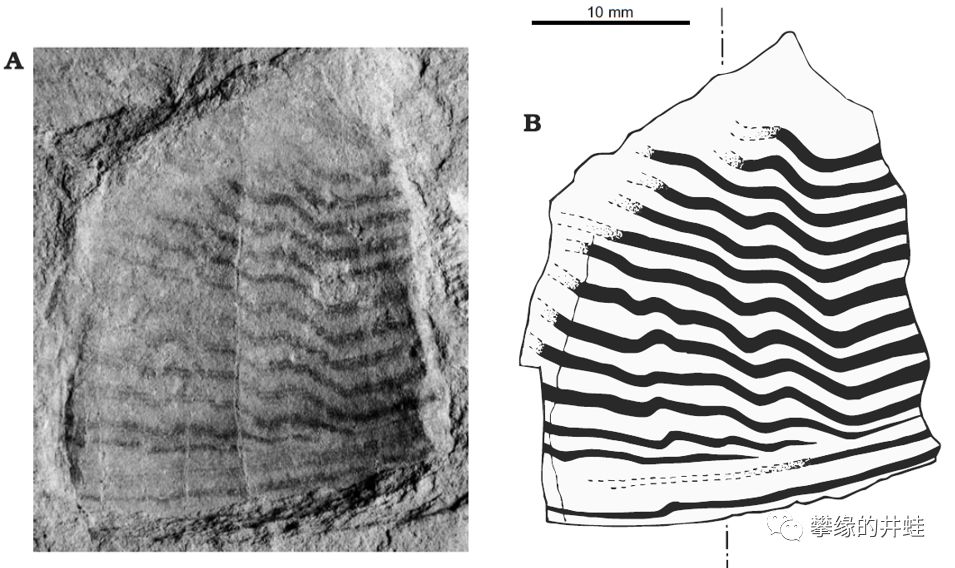

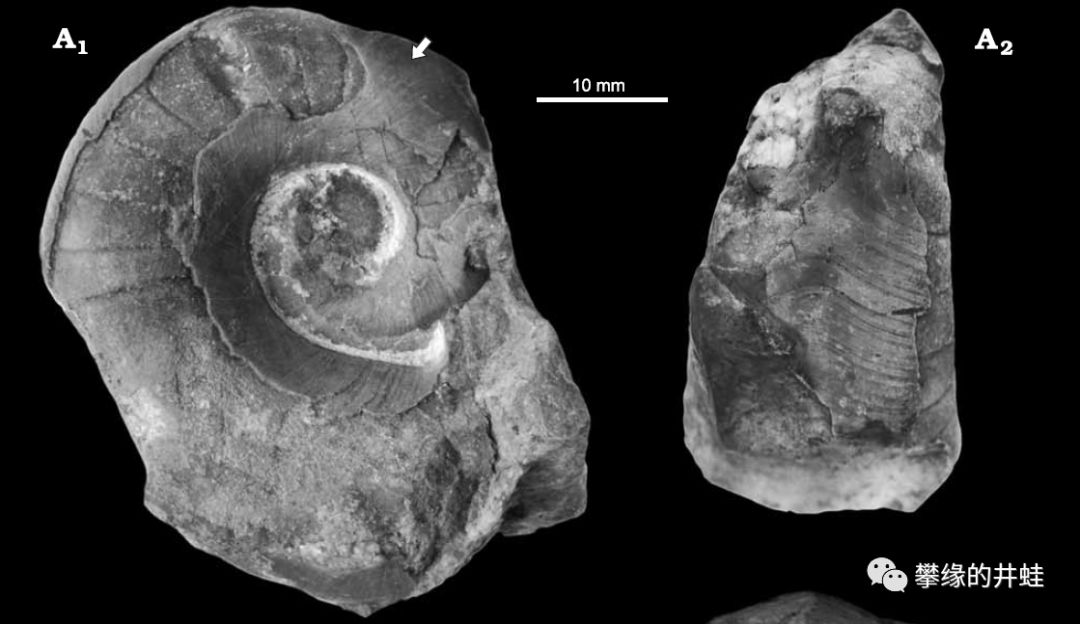
一些保存了表面纹饰的泥盆纪直角石和鹦鹉螺化石。发现地:捷克。看来头足动物外壳上一直都是这种波浪条纹,没有太大变化。图片来源自[9]。
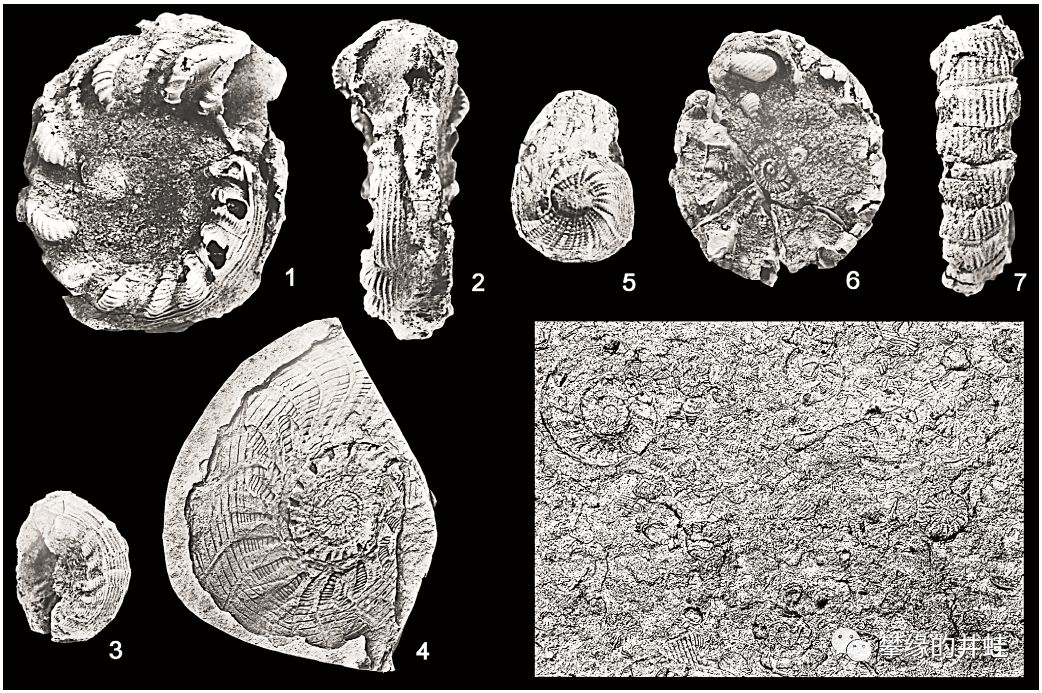
南京发现的二叠纪菊石Nodogastrioceras discum。贝壳表面有发达的筋肋结构,像拧成麻花一样。图片来源自[10]。
头足动物身上的坚硬结构还有喙(Beak)和齿舌(Radula),是和有颌鱼类抗衡的利器。不过这些结构不是钙盐,而是硬化的角蛋白,形成化石的概率比贝壳低得多,但也有不少发现。

一些内部带有喙结构的晚泥盆世菊石化石。螺壳内部的黑色方块就是喙的印痕。发现地:摩洛哥小阿特拉斯山(Anti-Atlas)地区。图片来源自[11],标尺长度1厘米。

也有和贝壳分离,单独发现的头足类喙化石。上图化石来自晚泥盆世,发现地:摩洛哥小阿特拉斯山(Anti-Atlas)地区。图片来源自[11],标尺长度1厘米。
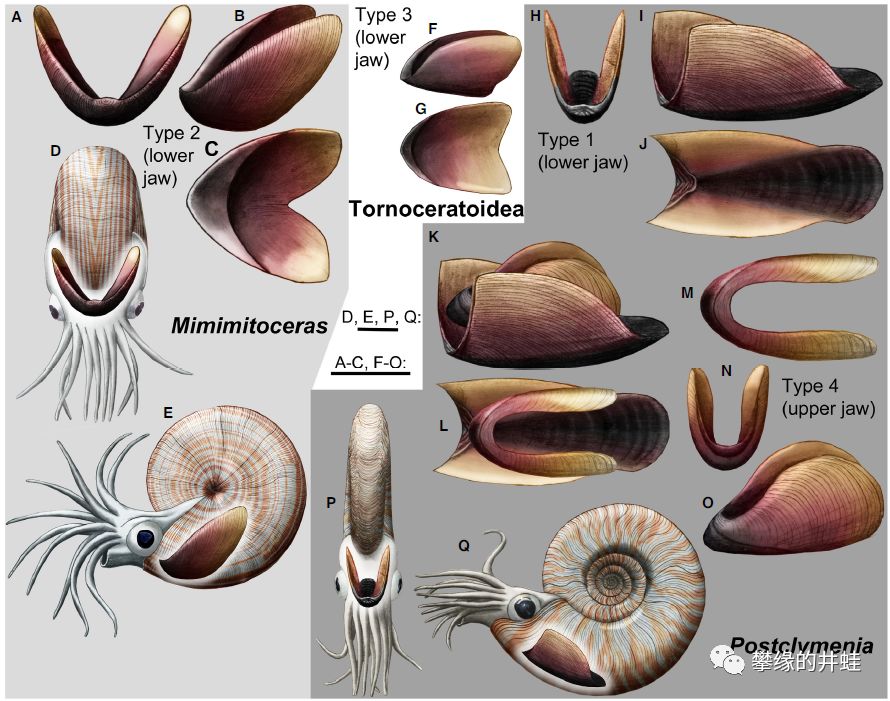
一些泥盆纪菊石复原图,标出喙(主要是下瓣)的位置和大小。图片来源自[11],标尺长度1厘米。
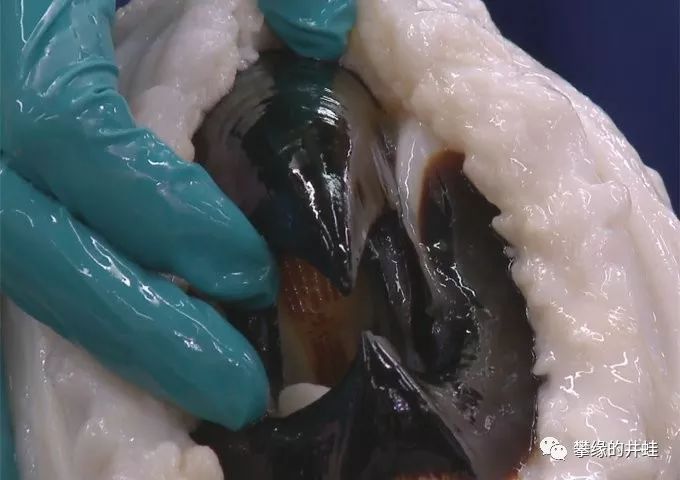
现代巨型鱿鱼的大口。鹦鹉嘴一样的喙可以轻易撕碎猎物。在口腔内侧,还可以看到成排的齿舌。图片来源自网络。
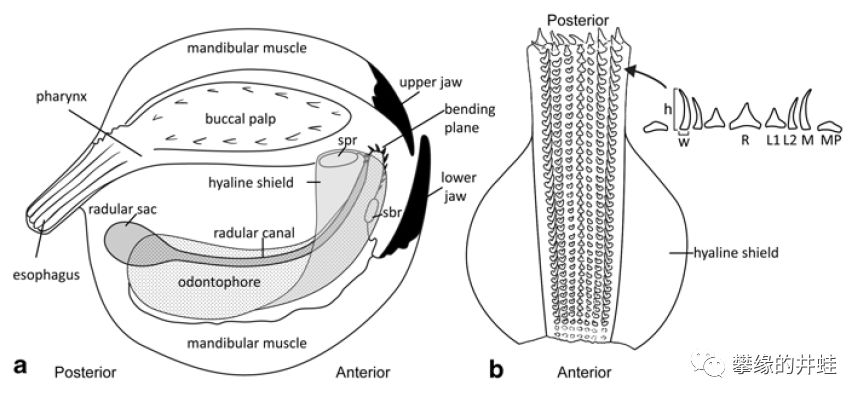
头足动物的进食构造。它们从刮食海藻的软体老祖宗那里继承了布满细小倒钩的齿舌。像传送带一样,把喙咬下的肉块送入食道。图片来源自[12]。
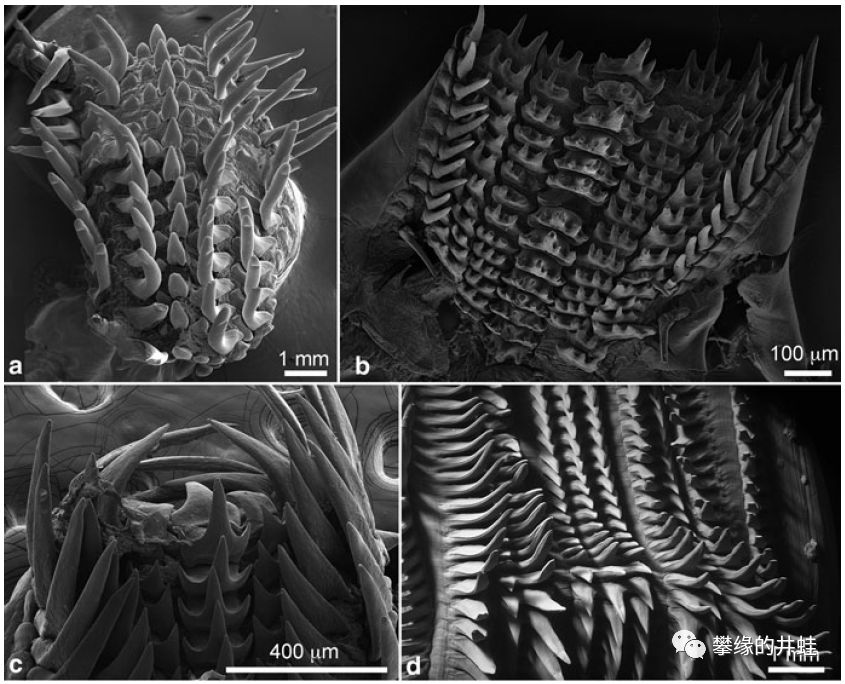
几类现代头足动物的齿舌。a:鹦鹉螺Nautilus pompilius;b:乍波章鱼Japetella diaphana;c:枪乌贼Loligo pealei;d:洪堡鱿鱼Dosidicus gigas。图片来源自[12]。

早二叠世菊石齿舌的印痕。和上图中现代头足动物齿舌的标尺对比一下,这个菊石体型相当大。产地:乌拉圭。图片来源自[12]。
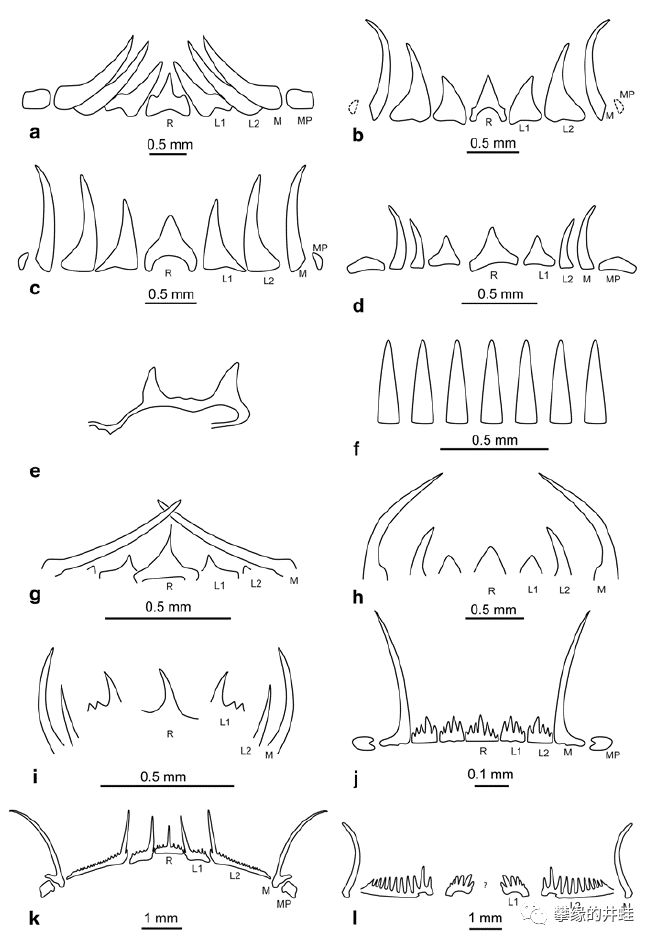
齿舌的形状是鉴别头足动物的重要依据。上图是几种不同菊石的齿舌倒钩样式。图片来源自[12]。
本回已经很长了。剩下的篇幅,再介绍一种奇特的头足动物:Sphooceras truncatum。S. struncatum生活在志留纪,广泛分布在当时的浅海,在世界各地都有发现,化石资料非常丰富。研究者在它的化石上找到了许多有意思的东西。

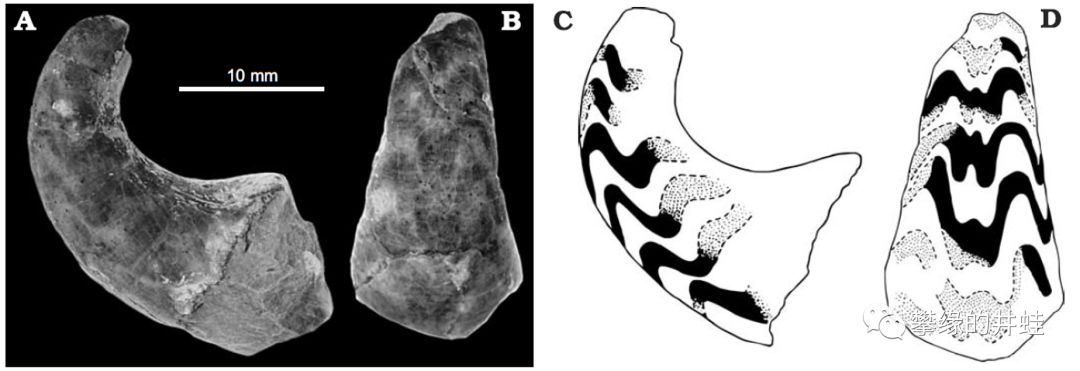
一些S. truncatum贝壳化石。产地:英国拉德洛(Ludlow)。它的贝壳结构接近直角石类,但是没有尖锐的壳顶,看上去圆滚滚的像一根香肠。图片来源自[13],标尺长度30毫米。

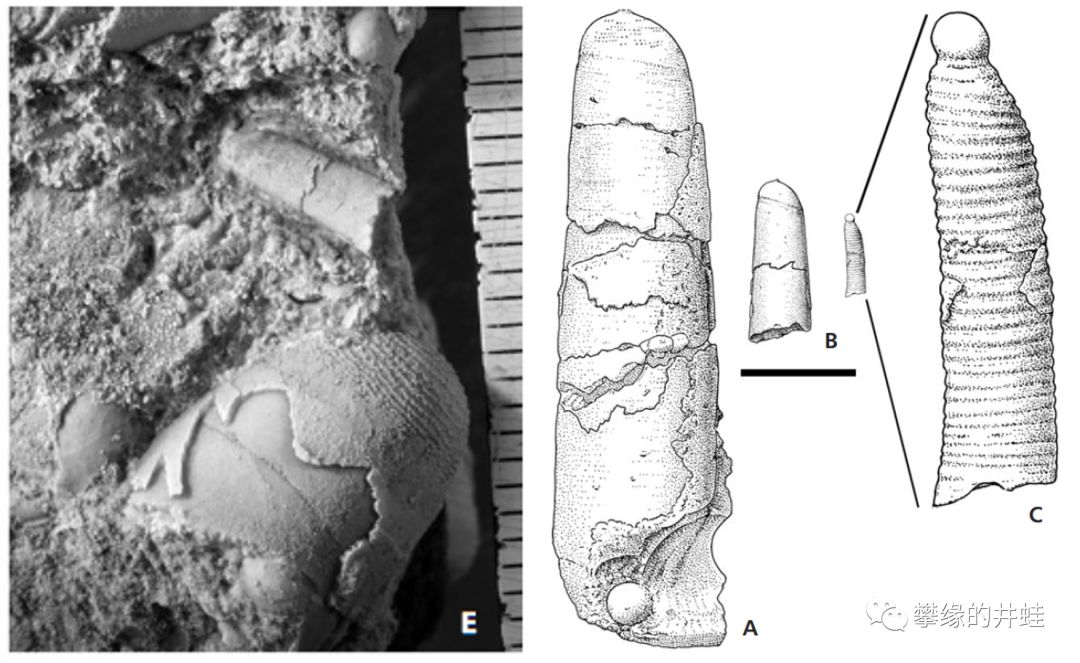
S. struncatum贝壳化石的截面,暴露出内部的腹管,隔板和气室。可以看到,气室在壳顶没有渐变,而是突然消失了。产地:英国拉德洛。图片来源自[13]。

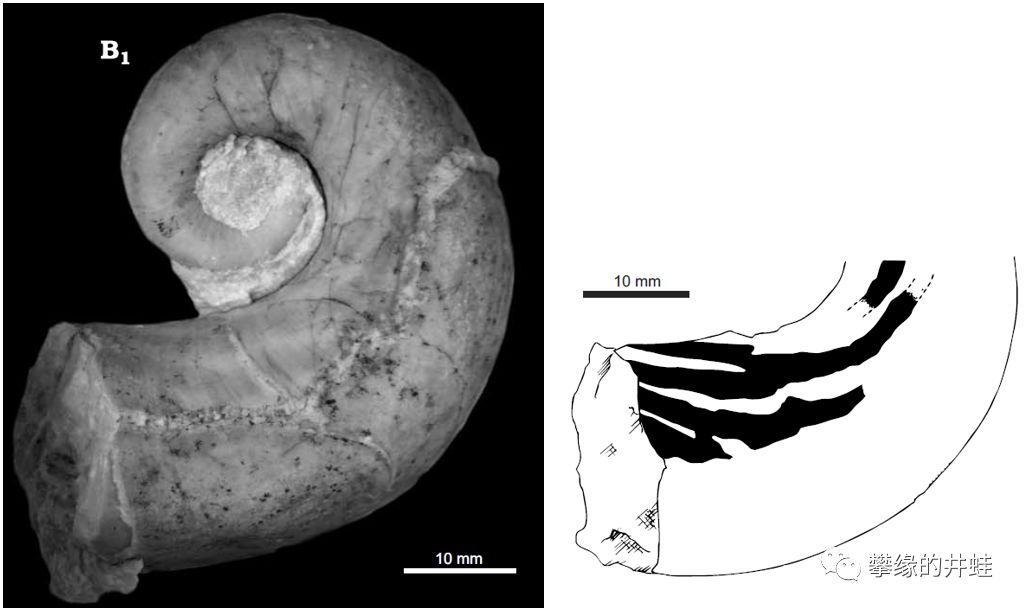
这些也是S. truncatum的贝壳化石。但是形状和上面的弯曲不一样。这是动物幼年时生长的部分,顶端的小圆包是刚孵化时长出的第一个螺壳(胚壳Protochonch)。但是从来没有在S. truncatum成体的化石上发现这个部分——它们在生长的过程中自然脱落了。图片来源自[13],标尺长度2毫米。
这是S. truncatum生长过程中一个很有意思的现象:贝壳尖端的部分会定期脱落(反正是中空的),只留下靠近体室的几个气室(成体是4个)。这大概和卷成螺旋形一样,也是一种避免壳体过长影响运动的方法。
S. struncatum生长早期贝壳(尖小)和成体贝壳(圆钝)对比。图片来源自[13],标尺长度5毫米。


有趣的是,一种现代蜗牛——断头钻头螺Rumina decollata也会把壳尖脱落掉,然后重新分泌钙质封闭壳顶。这种蜗牛在上海的城市绿化带里很常见,井蛙不止一次观察到它们聚在一堆大嚼狗屎。它们的壳尖非常脆弱,保存起来挺困难的。
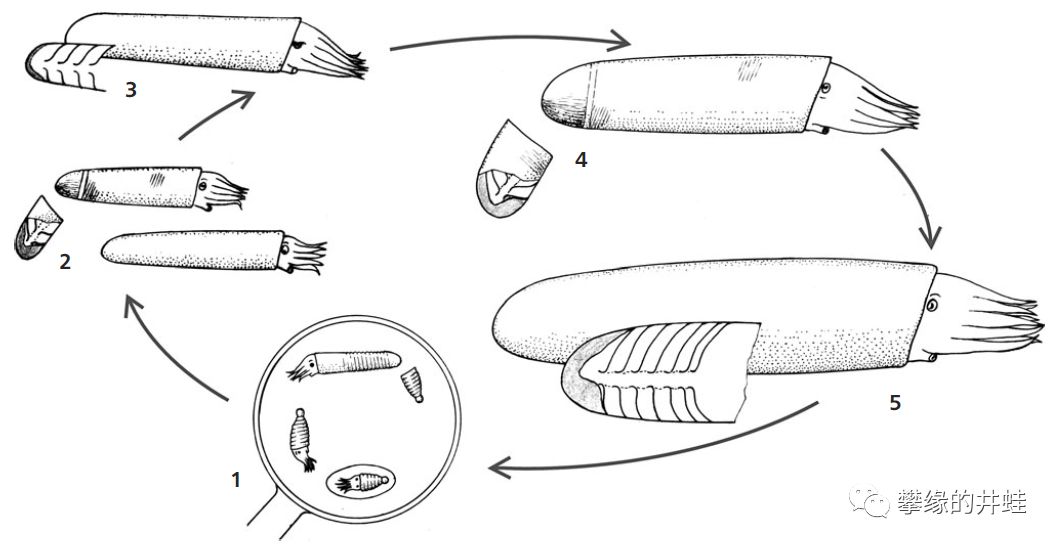
S. struncatum从孵化到成体的发育过程。孵化后,随着身体的成长,壳尖不断脱落,断裂的壳顶重新被钙质封闭,变成圆圆的钝头。S. truncatum的另一个奇特之处是:从压成体开始,外套膜就从壳口翻出来包裹住外壳,而成体的贝壳是全部埋在软体内部的(化石贝壳表面的特征证明了这一点)。这使它们看上去更像一根肉肠。图片来源自[13]。

S. struncatum成体的复原图。图片来源自网络。
S. struncatum的发现表明,就像贝壳卷曲成螺旋形一样,头足动物的演化史上也多次出现贝壳退化进入体内,软体完全暴露在外面的类型。属于直角石类的S. truncatum可能是最早的探索者,但它显然没能走到最后,只在志留纪昙花一现后就永远消失了,也没有后裔留下。在这条演化道路上,真正取得成功的是蛸形类。在身披重甲的头足类步入消亡后,那些赤膊上阵的触手怪们依然在世界各地的海洋中挥舞着它们的触手,仿佛胜利的旗帜。关于蛸形类的故事,请听下回分解。
地球名片
生物分类:动物界-软体动物门-头足纲-鹦鹉螺亚纲-直角石目
存在时间:奥陶纪早期至三叠纪末期
现存种类:无
化石种类:不明
生活环境:海洋
代表特征:圆锥形贝壳,腹管位于贝壳中心,简单缝合线
代表种类:契林角石,震旦角石,Sphooceras truncatum等
[1] Shuichi Shigeno, Sasaki Takenori, Sigurd vonBoletzky, The origins of cephalopod body plans: A geometrical and developmentalbasis for the evolution of vertebrate-like organ systems. Cephalopods - Presentand Past, Tokai University Press, Tokyo, p. 23-34, 2010
[3] ED LANDING,BJÖRN KRÖGER, THE OLDEST CEPHALOPODS FROM EAST LAURENTIA. J. Paleont., 83(1),2009, pp. 123–127
[2] ChristianKlug, Björn Kröger, Jakob Vinther, et al., Ancestry, Origin and EarlyEvolution of Ammonoids. Ammonoid Paleobiology: Frommacroevolution to paleogeography, Geobiology 44, DOI10.1007/978-94-017-9633-0_1
[4] Pohle, A.& Klug, C. 2018: Body size of orthoconic cephalopods from the late Silurianand Devonian of the Anti-Atlas(Morocco). Lethaia, Vol. 51,pp. 126–148, DOI 10.1111/let.12234
[5] Lutz Koch, Stefan Voigt, CarstenBrauckmann, Nautiliden aus der Kluterthöhle (Ennepetal, Nordrhein-Westfalen),aus benachbarten Höhlen und weiteren Fundorten in Oberen Honsel-Schichten(Unter-Givetium). Geologie und Paläontologie in Westfalen, 90: xx-xc
[6]Lutz Koch, Ennepetal Stefan Voigt,Ennepetal Carsten Brauckmann, et al., Nautiliden-Funde aus der Kluterthöhle undder Heilenbecker Höhle (Ennepetal, Nordrhein-Westfalen) mit einem Beitrag zumLebensraum der Nautiliden in den Korallen-Stromatoporen-Riffen der OberenHonsel-Schichten (Unter-Givetium). Dortmunder Beitr. zur Landesk., naturwiss.Mitt., 48, 77–96, 2018
[8] Royal H. Mapes, Neil H. Landman,Christian Klug, Caught in the act? (Distraction sinking in ammonoidcephalopods). Swiss Journal of Palaeontology,https://doi.org/10.1007/s13358-018-0176-7
[7] Klug, C. and Korn, D. 2004. The origin of ammonoidlocomotion. ActaPalaeontologica Polonica 49(2): 235–242
[9] Turek,V. 2009. Colour patterns in Early Devonian cephalopods from the BarrandianArea: Taphonomy and taxonomy. Acta Palaeontologica Polonica 54 (3): 491–502.DOI: 10.4202/app.2007.0064
[10] ZURENZHOU, BIZARRE PERMIAN AMMONOID SUBFAMILY AULACOGASTRIOCERATINAE FROM SOUTHEASTCHINA. J. Paleont., 81(4), 2007, pp. 797–799
[11] CHRISTIANKLUG, LINDA FREY, DIETER KORN, et al., THE OLDEST GONDWANAN CEPHALOPODMANDIBLES (HANGENBERG BLACK SHALE, LATE DEVONIAN) AND THE MID-PALAEOZOIC RISEOF JAWS. Palaeontology, 2016, pp. 1–19
[12] Isabelle Kruta, Neil H. Landman,Kazushige Tanabe, Ammonoid Radula. Ammonoid Paleobiology: From anatomy toecology, 2015, DOI: 10.1007/978-94-017-9630-9_11
[13] TUREK, V., MANDA, Š., “Anendocochleate experiment” in the Silurian straight-shelled cephalopod Sphooceras.Bulletin of Geosciences 87(4), 767–813, DOI 10.3140/bull.geosci.1335