
直说吧,这回讲鳞鲵类(Microsauria),属于壳椎类中的一支,也是“古两栖类”。
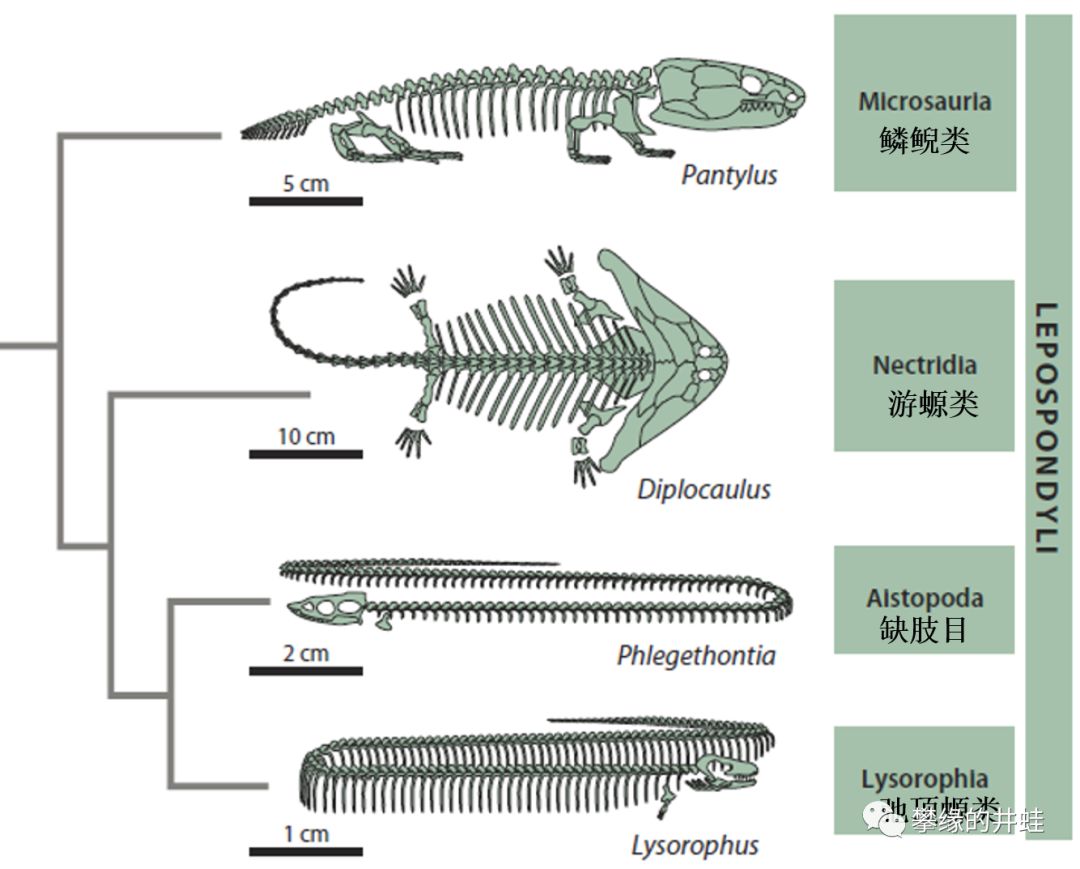
研究者曾经把壳椎类分为鳞鲵目,游螈目,缺肢目,还有弛顶螈类。不过新的观点认为,这种分类方法并不符合系统发生学的原则。现在“鳞鲵类”更多地是一种并不严格的惯称,就像“鱼类”,“两栖类”,“爬行类”一样。

一种比较新的壳椎类演化关系图。除了缺肢目外,“鳞鲵目”,“游螈目”和“弛顶螈类”都不是单系群。尤其是“鳞鲵目”,基本就是个筐,凡是没有什么特点的壳椎类都被扫到里面了。这么多的种类,却找不到几个中文译名,可见它们的知名度实在不怎么样。图片来源自[1]。
关于缺肢目,弛顶螈类,还有游螈目的棒尾螈科和笠头螈科可以参看第一百九十二回Normal to Bizarre和第二百三十回螈怪,都是些奇形怪状的特化物种。和它们相比,鳞鲵类大体处在壳椎类的基干位置,身体没有发生很极端的特化。
所以鳞鲵类最大的特点就是——没特点。虽然在专业的研究者眼中,每一种生物都是独特的,但是对普通的爱好者来说,鳞鲵类体型小,样子普通,既不威猛,又不怪异,生活方式也没什么特别,放在各种四足怪兽横行的石炭纪和二叠纪,很难像异齿龙,杯鼻龙,锯齿螈,引螈,笠头螈那样,给人留下深刻的印象。是啊,帅卖怪坏哪一样都不占,让我怎么捧你?让观众怎么记住你?

一些鳞鲵类的复原图。A:Hyloplesion;B:Pantylus;C:Pelodosotis;D:Rhynchonkos。抱歉,我找不到它们的中文译名,还请大神们指点。能看出这几个货和蝾螈,蜥蜴有什么不一样吗?图片来源自Wikimedia。
但鳞鲵类又确实是壳椎类中种类最多,多样化程度最高的一群。把它们撇开不写,真有点说不过去。
所以井蛙在整理的资料的时候,心里默默地给了鳞鲵类一个评价,就是“尴尬”。就连这篇文章,也像是在和读者朋友们有一搭没一搭的强行尬聊。
那就聊到哪儿算哪儿吧。
“Microsaur”直译应该是“微龙”或者“微蜥”。日语翻译成“細竜目”,也很贴切。不知为什么中文称为“鳞鲵”,也许是更强调“两栖动物”的身份吧。顾名思义,它们的体表是覆盖有鳞片的,但这根本称不上特点。因为绝大多数古生代的两栖动物,无论离片椎类还是壳椎类,身上都有鳞片(参见第二百二十九回蟾祖)。
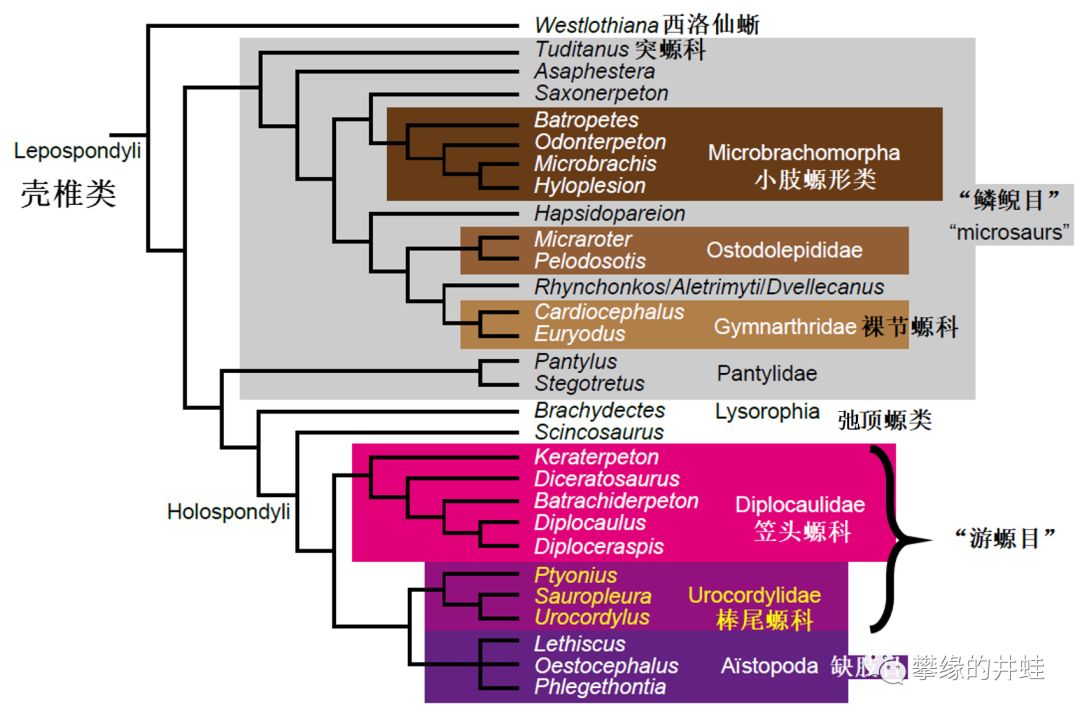
一种石炭纪晚期鳞鲵类:小肢螈Microbrachis pelikani的鳞片化石。A-D是单个鳞片,E是鳞片在皮肤上排列成行。图片来源自[2],标尺长度1毫米。

捷克发现的晚石炭世鳞鲵类Sparodus validus(裸节螈科)化石。这只动物在石化的过程中被压扁挤碎了,但还能看出一些部分的形状,比如左上方的肢骨,中间的脊柱和肋骨,还有右下方的颅骨。散布在骨骼四周的那些圆圆的小东西就是它的鳞片。图片来源自[3]。
研究者曾经认为全部或者至少一部分滑体两栖动物(无足目)起源自鳞鲵类。但是分子生物学和过渡类型化石的证据都显示,离片椎类才是现代两栖动物的祖先。于是鳞鲵类就沦为了一个拔丝无后的演化旁支,彻底断了香火。
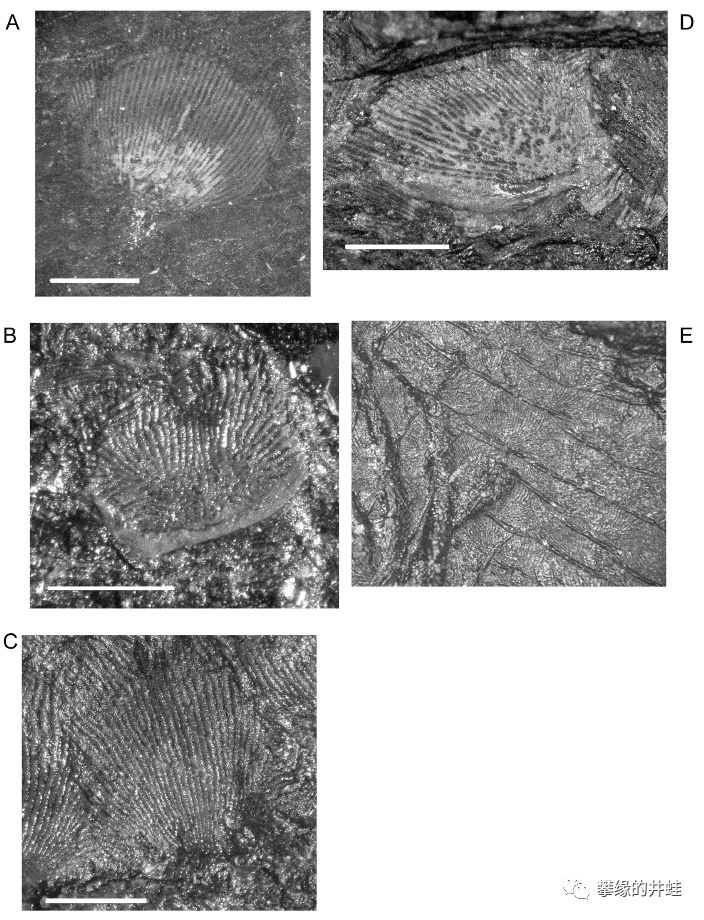
几种曾经被认为和滑体两栖类起源关系密切的古两栖动物。A–C:Brachydectes(弛顶螈类)。D–F:Rhynchonkos(鳞鲵类)。G–I:Doleserpeton(离片椎类—双疏螈类)。在第二百二十九回蟾祖里介绍过,目前的证据显示蛙形类(无尾目和有尾目)起源自双疏螈类的一支,无足目也和鳞鲵类没有关系。图片来源自[4]。
鳞鲵类的种类不少,生活方式也挺多样,有的终生水栖,有的成体陆栖,也有的在地下穴居。留下的化石也挺丰富的,其中不乏保存完整的珍品。但是因为知名度太低,不被关注,在网上能找到的化石照片和复原图都不多。

俄克拉荷马州Richards Spur发现的鳞鲵类Cardiocephalus peabodyi(裸节螈科)的完整化石。它生活在二叠纪早期,体长大约20厘米,脊柱延长,四肢纤弱,表明是高度水栖的动物。图片来源自网络。
再简单介绍一些比较重要的鳞鲵类吧。
Pantylus
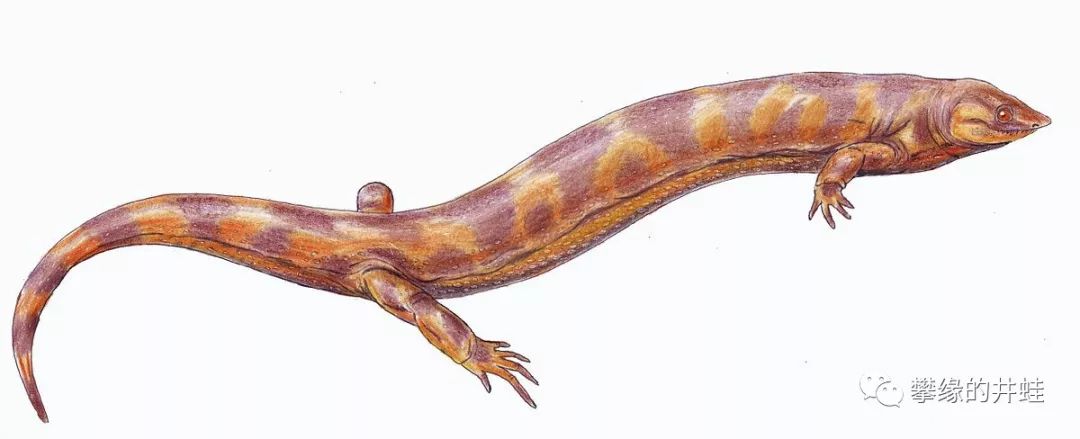
是Pantylidae科下的一属,只包含一个种:Pantylus cordatus。化石发现于北美的早二叠世地层,成体体长大约25厘米,身体结构有明显的陆栖特征:有着不成比例的大头,身体,四肢和尾巴全都又短又粗,有点像拔了毛的竹鼠。
Pantylus cordatus复原图,来源自Wikipedia。小别致长得真东西啊。

盘起来的话,感觉大概是这样的?图片来源自网络。

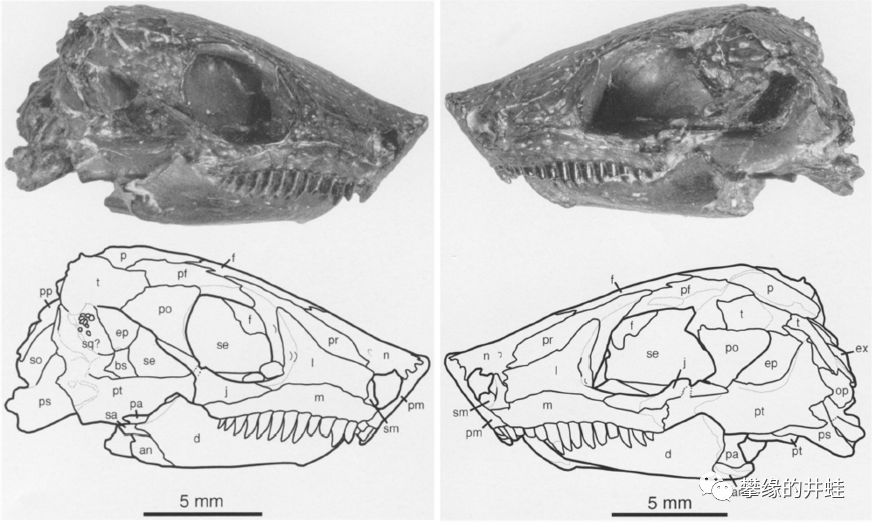
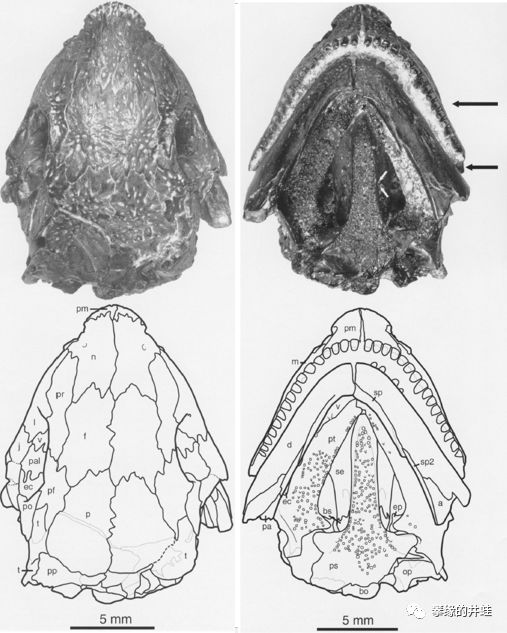
Pantylus cordatus头骨化石。它的体型不像能快速奔跑的样子;脑袋坚实笨重,牙齿大而钝,可能主要捕食蜗牛,昆虫,蛛形类等长有硬壳的无脊椎动物,应该也不会拒绝蠕虫,羊膜类的卵和幼体,还有腐肉。图片来源自[5]。
在奇葩云集的壳椎类中,鳞鲵类大都顶着一张路人脸。所以唯一还算有点特色的P. cordatus经常被拿来充门面。图片来源自[6]。
Ostodolepidae
这是一个科,和Pantylus亲缘关系很近,也是高度陆栖的,最大的种类有60厘米长。在发现之初,曾经被当做原始的羊膜动物。它们的头短而尖,躯干很长,尾巴较短,四肢粗壮有力,生活方式可能像今天的一些穴居石龙子。
Ostodolepidae科Pelodosotis属动物的复原图,来源自Wikipedia。
一种早二叠世Ostodolepidae科动物Nannaroter mckinziei的头骨化石。发现地:俄克拉荷马州Richards Spur。这个科的动物都有一个上翘的小“猪鼻”,挺可爱的。图片来源自[7]。
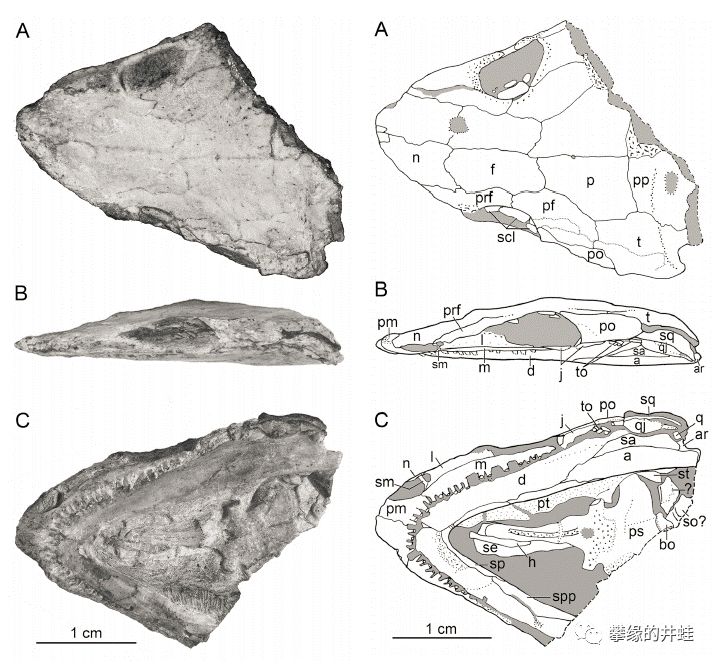
德国中部地区发现的早二叠世Ostodolepidae科动物Tambaroter carrolli头骨化石。从复原图上看,也长了个小小的猪鼻子。图片来源自[8]。
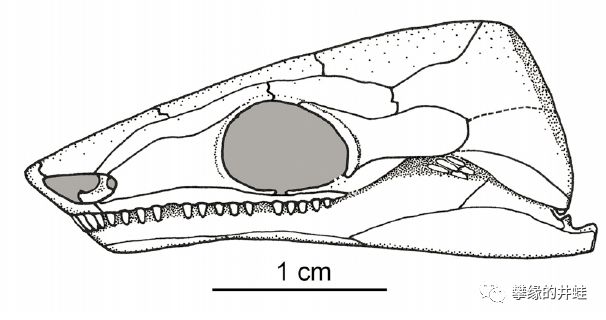
一种早二叠世Ostodolepidae科动物Brevispinatus Williston,化石近乎完整。发现地:德克萨斯州咖啡溪(Coffee Creek)。图片来源自[9]。
小肢螈科(Microbrachidae)
是一些高度水栖的小型(成体不到20厘米)动物,许多种类终生保留外鳃。生活在淡水环境中的动物被掩埋石化的几率要高一些,所以小肢螈科留下了大量化石资料,特别是Microbrachis pelikani和Hyloplesion longicostatum,包括各个发育阶段的幼体,是研究和了解最为深入的鳞鲵类。然而并没有什么卵用,在大众当中的知名度和辨识度极底。

Hyloplesion longicostatum(a)和Microbrachis pelikani(c)完整化石。发现地:捷克;生存年代:石炭纪晚期。它们有多达40块躯干脊柱,像一条被抻长了的蝾螈。图片来自[10],标尺长度:a,c,1毫米;b,2毫米。
Microbrachispelikani复原图。注意一个小细节:它的前肢有5个脚趾。绝大多数“两栖动物”,包括离片椎类,壳椎类和滑体两栖类,前肢都只有4个脚趾。图片来源自网络。
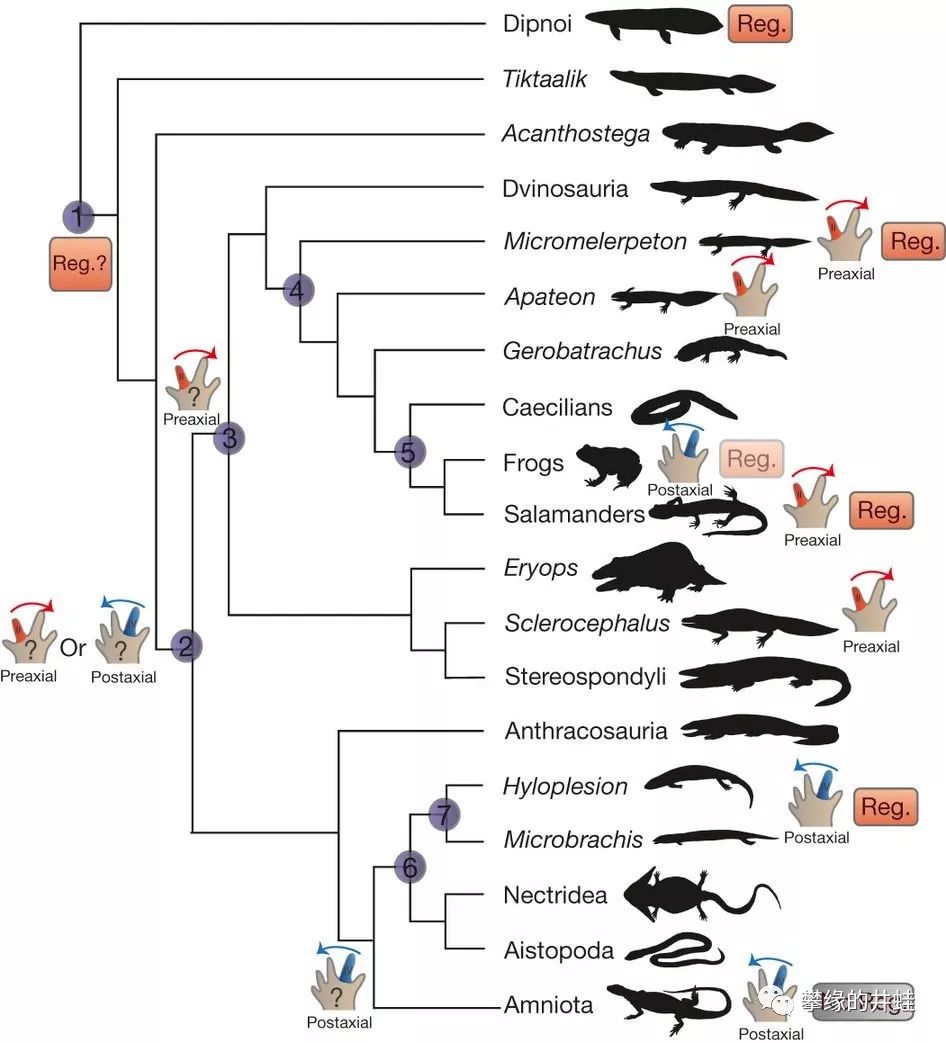
一些灭绝和现存四足动物前肢脚趾的数量对比。注意只有小肢螈科(7)和羊膜类(最下)保留了5根脚趾。这也是壳椎类和羊膜动物亲缘关系的一个证明。蛙的旁边也画了5根脚趾,因为无尾目在蝌蚪阶段末期,前肢肢芽上也是有5根脚趾的,但在变态完成后有一根退化消失了。图片来自[10]。
前面说过,小肢螈类留下了不同阶段幼体的化石。它们展示了壳椎类和离片椎类发育过程的巨大差异。
有朋友问锯齿螈的蝌蚪有多大。答案可能出乎意料:很小,就和你平时见到的蝌蚪差不多大。因为非羊膜卵的限制,无论最终的成体有多大,刚刚孵化的幼体都很小,发育很不完全。
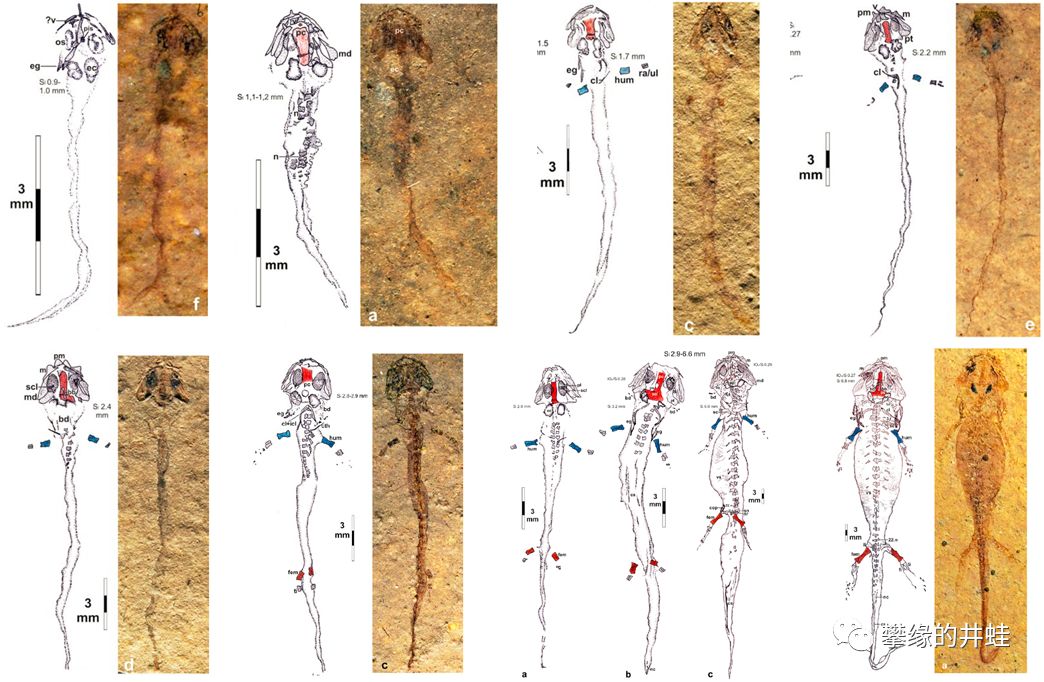
一种小型离片椎类幻螈Apateon的早期发育过程。刚刚孵化的幼体没有四肢,连脊椎都没有骨化,是真正的“蝌蚪”。随后先萌发前肢,然后萌发后肢(这个顺序和现代有尾目一样),摆脱蝌蚪形态,成为亚成体(幼鲵)。图片来源自[11]。
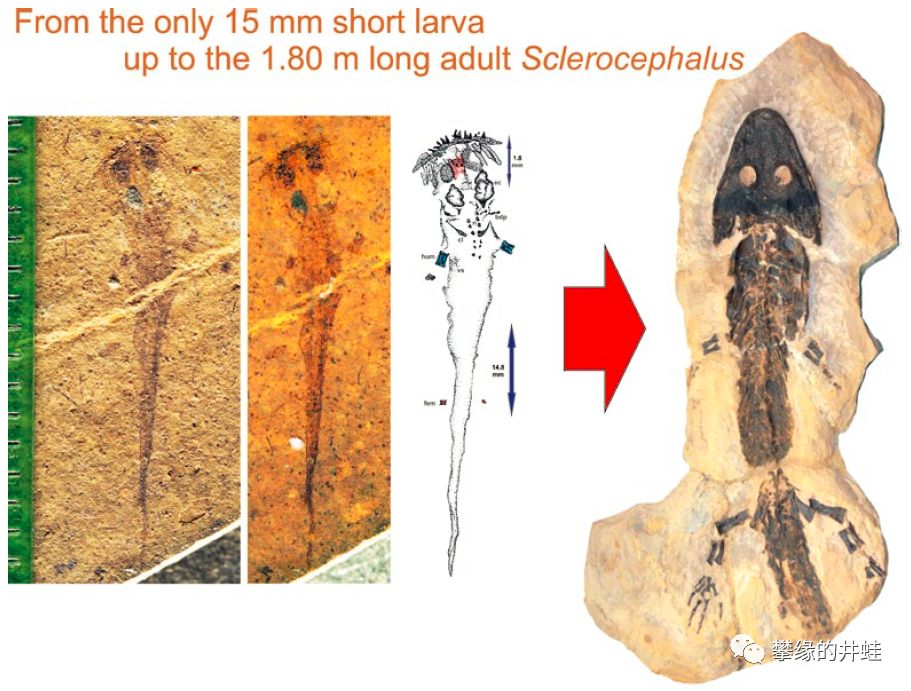
大型离片椎类Sclerocephalus,刚孵化的蝌蚪也只有1.5厘米。它的幼鲵阶段会比较长,最后发育成1.8米长的大家伙。比它更加巨大的始鲵类也是一样。图片来源自[11]。
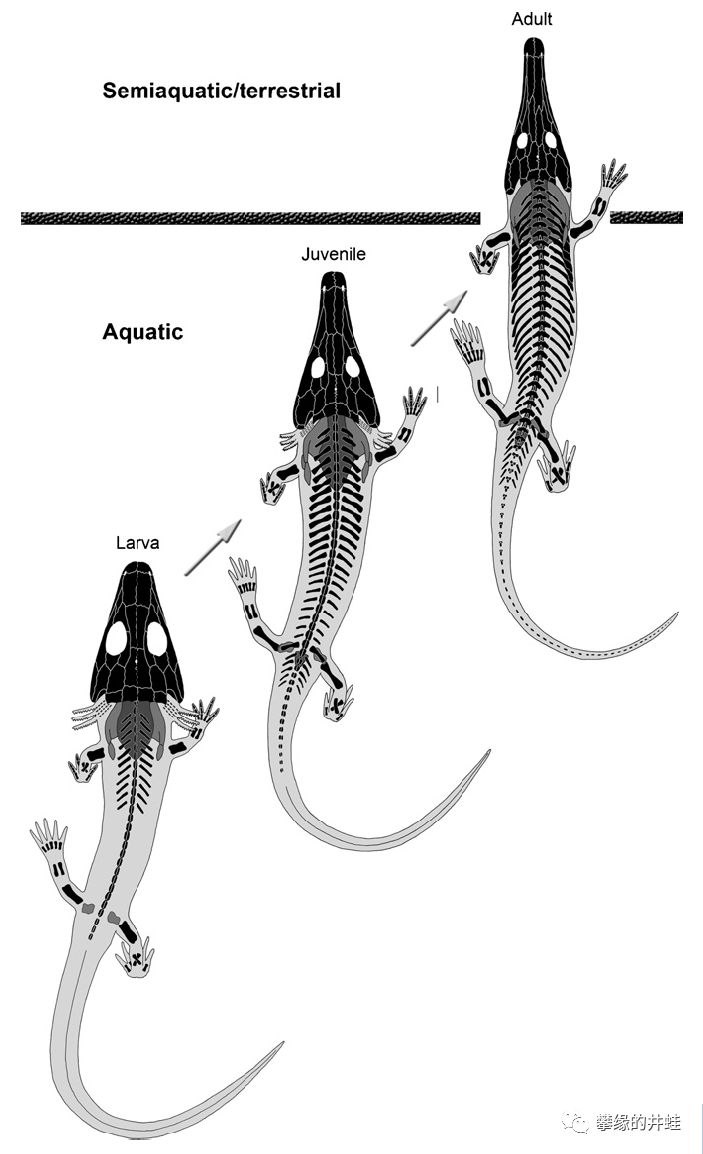
大型离片椎类从幼鲵到成体的发育过程。外鳃萎缩消失,头骨延伸变形,同时伴随的还有脊柱,肋骨和肢骨的钙化成型。图片来源自[12]。
所以离片椎类的脊柱是在发育后期骨化的,晚于肢骨。离片椎类幼体的化石上往往观察不到成型的脊椎骨。
而壳椎类恰恰相反,目前发现的最小的壳椎类幼体,都有骨化完全的脊椎和肢骨。换言之,壳椎类的幼体发育非常完善,完全没有软趴趴没四肢的“蝌蚪”阶段。
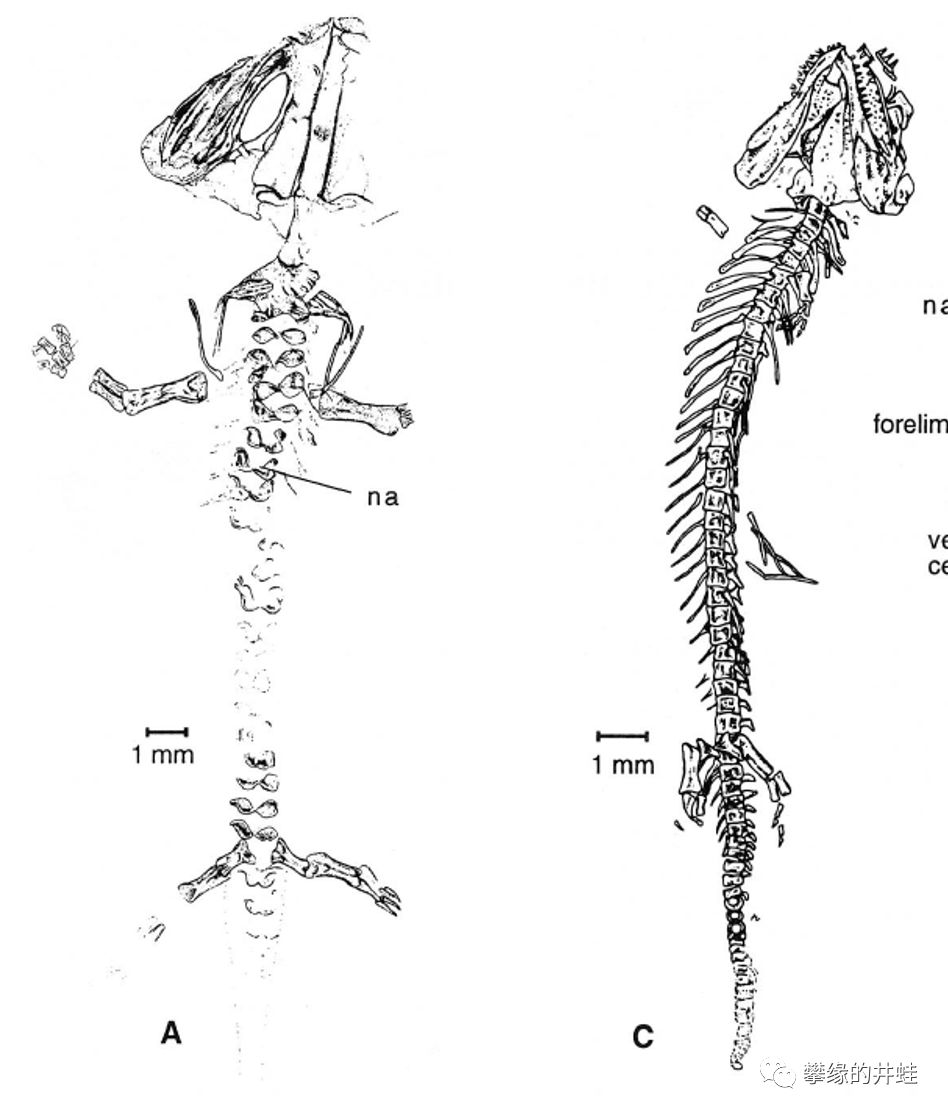
体型接近的小型离片椎类(左:鳃龙科的Branchiosaurus salamandroides)和壳椎类(右:小肢螈科的Hyloplesion longicostatum)幼体骨骼对比。壳椎类幼体的脊柱明显比离片椎类幼体发育完善得多。图片来源自[13]。
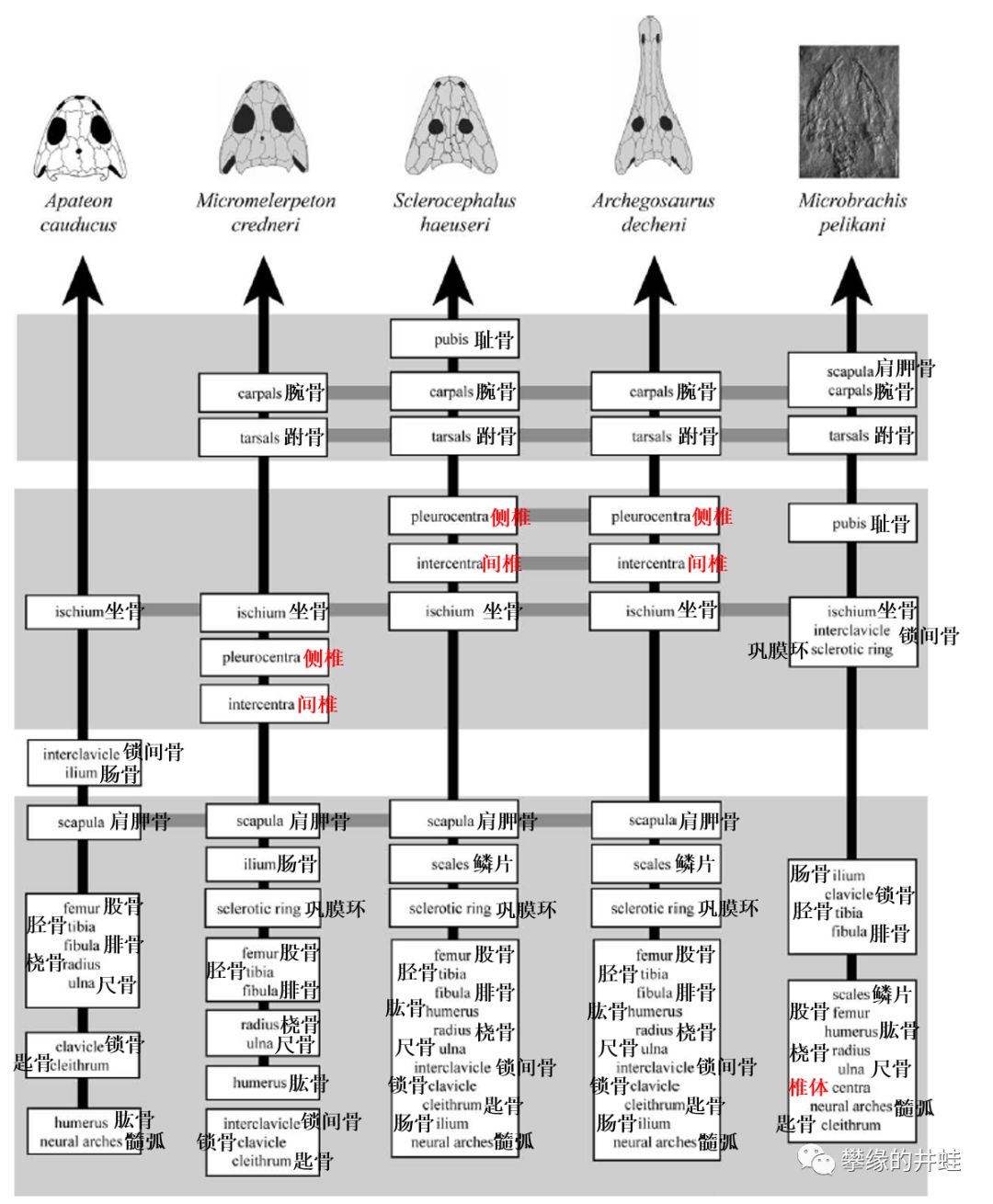
离片椎类(左四列)和壳椎类(最右列)骨骼发育顺序对比。尤其是脊椎椎体(红字)的骨化时间差别明显。图片来源自[14]。
一般来说,发育如此完全的幼体意味着更大的卵,和更长的孵化时间,比起两栖类,这些特征更进步一些,更接近羊膜动物。那么壳椎类到底是怎样繁殖的?它们是不是会产下某种“半羊膜卵”?这些问题,大概永远不会有答案了。
现在看来,鳞鲵类始终处在一个非常尴尬的地位,在水中竞争不过离片椎类,陆地又被真正的羊膜类先行一步占领了。它们只能憋憋屈屈跑跑龙套,填补一些边边角角的生态位。
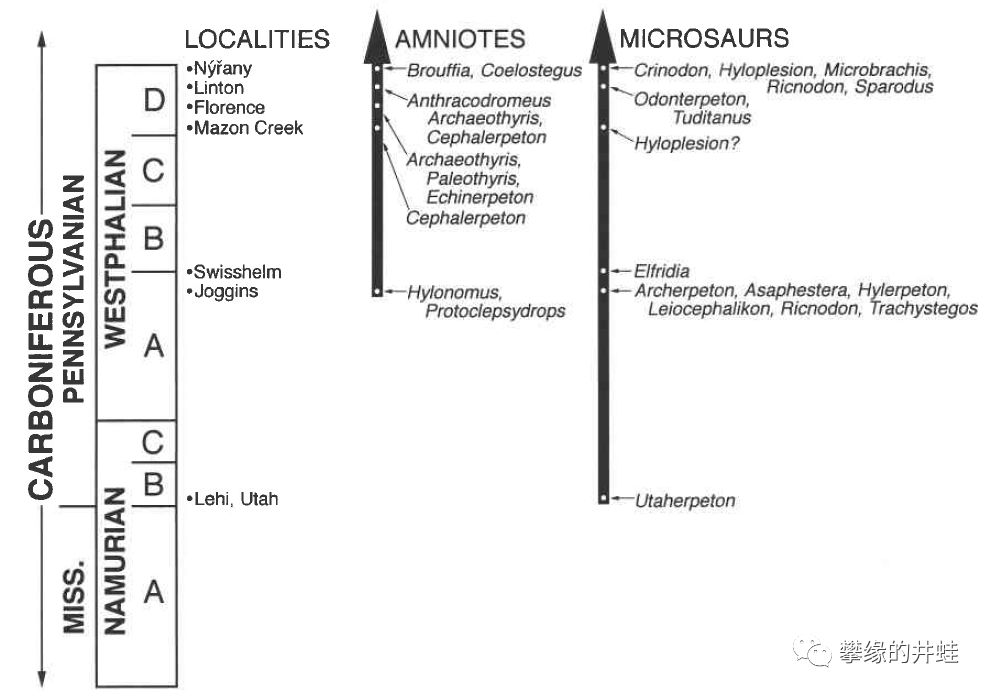
早期羊膜动物(左箭头)和鳞鲵类(右箭头)的生存时间和化石产地,图片来源自[15]。目前发现的最早的鳞鲵Utaherpeton的生存时间要早于林蜥Hylonomus和Protoclepsydrops(可能是最早的合弓纲动物)。“明明是我先的啊!”
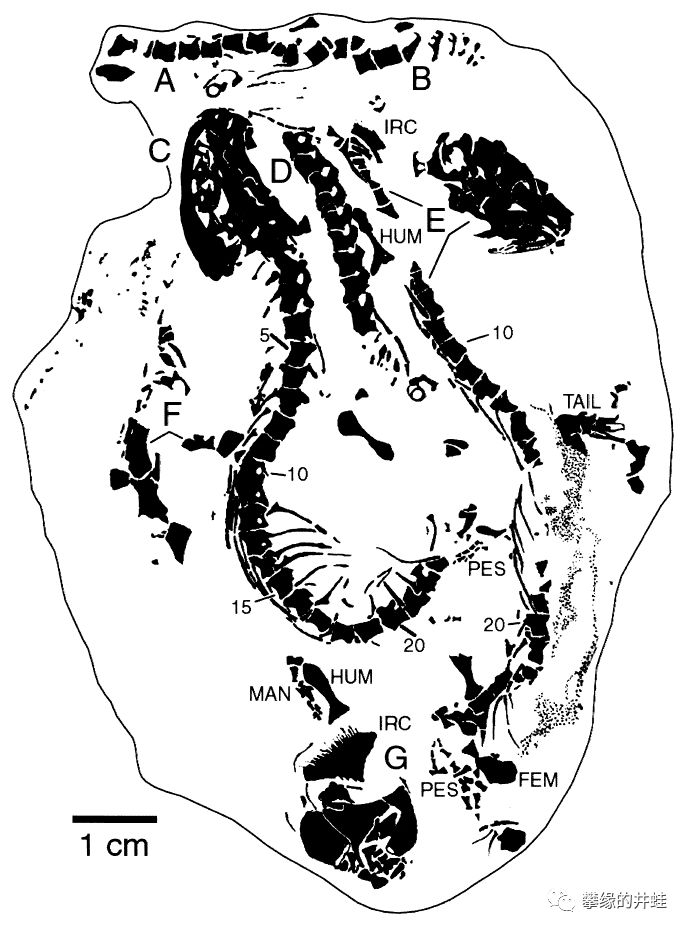
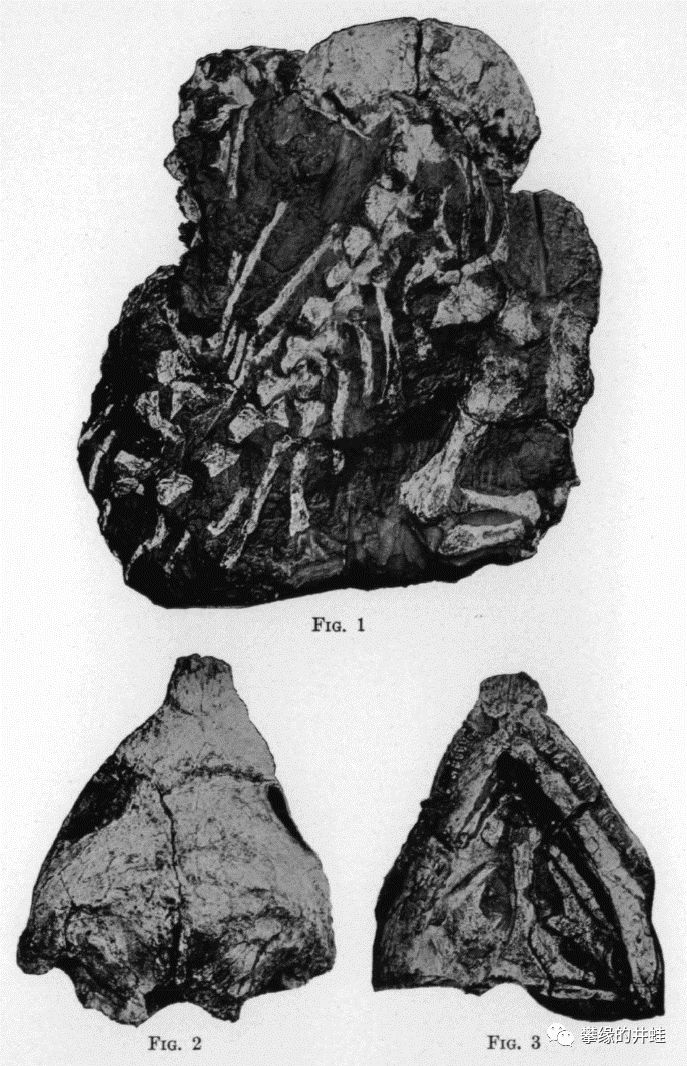
一块早石炭世鳞鲵类化石(未查到学名),来自伊利诺伊州戈尔维尔(Goreville)。这块化石上有至少8只鳞鲵类挤在一起。陆栖鳞鲵类和羊膜动物的身体结构非常接近,连专业的古生物学者都经常搞混。它们的生存能力应该不会弱于林蜥,古窗龙和始祖单弓兽。然而不知为什么,鳞鲵类始终没能成为演化舞台上的主角。图片来源自[16]。
鳞鲵类的演化努力在二叠纪中期画上了句号,甚至没能见识超级大灭绝的威力,也不知道是幸运还是不幸。之后不久,最后的壳椎类(笠头螈)也没能逃过一劫,四足动物演化史上这个并不算成功的分支,彻底沉寂在漫漫历史长河中。
地球名片
生物分类:动物界-脊索动物门-脊椎动物亚门-四足形类-壳椎类-鳞鲵类
存在时间:石炭纪早期至二叠纪末
现存种类:无
化石种类:超过30个属,未找到完整种类统计
生活环境:淡水,陆地
代表特征:体型小到中型,很可能是多系群,包括所有被游螈类,缺肢类,弛顶螈类排除在外的壳椎类
代表种类:Hyloplesion,Pantylus,Pelodosotis,Rhynchonkos,Utaherpeton,Hyloplesion,Microbrachis,等等
[1] DavidMarjanović, Michel Laurin, Reproducibility in phylogenetics: reevaluation ofthe largest published morphological data matrix for phylogenetic analysis ofPaleozoic limbed vertebrates. DOI: 10.7287/peerj.preprints.1596v3
[2] Olori JC (2015), Skeletal Morphogenesisof Microbrachis and Hyloplesion (Tetrapoda: Lepospondyli), and Implications forthe Developmental Patterns of Extinct, Early Tetrapods. PLoS ONE 10(6):e0128333, doi:10.1371/journal.pone.0128333
[3] Marjanović D, Laurin M. 2019. Phylogeny of Paleozoic limbed vertebratesreassessed through revision and expansion of the largest published relevant datamatrix. PeerJ 6:e5565, DOI 10.7717/peerj.5565
[4] TROND SIGURDSEN, DAVID M. GREEN, Theorigin of modern amphibians: a re-evaluation. Zoological Journal of the LinneanSociety, 2011, 162, 457–469
[5] http://fossils.valdosta.edu/fossil_pages/fossils_per/a10.html
[6] Rainer R. Schoch, Evolution of LifeCycles in Early Amphibians. Annu. Rev. Earth Planet. Sci. 2009. 37: 135–62, doi: 10.1146/annurev.earth.031208.100113
[7] Jason S. Anderson, Diane Scott, RobertR. Reisz, Nannaroter mckinziei, a New Ostodolepid 'Microsaur' (Tetrapoda,Lepospondyli, Recumbirostra) from the Early Permian of Richards Spur (Ft.Sill), Oklahoma. Journal of Vertebrate Paleontology, Vol. 29, No. 2 (Jun. 12,2009), pp. 379-388
[8] Amy C. Henrici, Thomas Martens, David SBerman & Stuart S. Sumida (2011): An ostodolepid ‘microsaur’(Lepospondyli)from the Lower Permian Tambach Formation of central Germany, Journal ofVertebrate Paleontology, 31:5, 997-1004,http://dx.doi.org/10.1080/02724634.2011.596601
[9] E. C. Case, Description of a NearlyComplete Skeleton of Ostodolepis Brevispinatus Williston. Contributions fromthe Museum of Paleontology – University of Michigan, Vol. III, No. 5, pp.81-107, 1929
[10] Nadia Fröbisch, Constanze Bickelmann, Jennifer C. Olori, et al., Deep time evolution of regeneration and preaxialpolarity in tetrapod limb development. October 2015, Nature 527(7577), DOI:10.1038/nature15397
[11] Ralf Werneburg, Schleusingen, Earliest‘nursery ground’ of temnospondyl amphibians in the Permian. Semana, Band 32,Seiten 3-42, 2017
[12] Nadia B. Frobischa, Jennifer C. Olori,Rainer R. Schochc, et al., Amphibian development in the fossil record. Seminarsin Cell & Developmental Biology, 21 (2010,) 424–431
[13] S. COTE, R. CARROLL, R. CLOUTIER, etal., VERTEBRAL DEVELOPMENT IN THE DEVONIAN SARCOPTERYGIAN FISH EUSTHENOPTERONFOORDI AND THE POLARITY OF VERTEBRAL EVOLUTION IN NON-AMNIOTE TETRAPODS.Journal of Vertebrate Paleontology, 22(3): 487–502, September 2002
[14] JenniferC. Olori, Ontogenetic sequence reconstruction and sequence polymorphism inextinct
taxa: an example using early tetrapods(Tetrapoda: Lepospondyli). Paleobiology, 39(3), 2013, pp. 400–428, DOI:10.1666/12031
[15] Robert Reisz, Sean Patrick Modesto,Archerpeton anthracos from the Joggins Formation of Nova Scotia: a microsaur,not a reptile. Canadian Journal of Earth Sciences, February 1996, DOI:10.1139/e96-053
[16] R. Eric Lombard, John R. Bolt, AMicrosaur from the Mississippian of Illinois and a Standard Format forMorphological Characters. Journal of Paleontology, Vol. 73, No. 5 (Sep., 1999),pp. 908-923