
祝所有的小朋友们六一儿童节快乐,今天我们要讲一个很久很久很久很久以前的故事。
当某种原始真核生命演化出以下技能时,有性生殖便正式出现了:
1.两个同类细胞可以发生融合,重组遗传物质;
2.融合后的新细胞可以发生减数分裂(Meiosis),平均分配遗传物质;
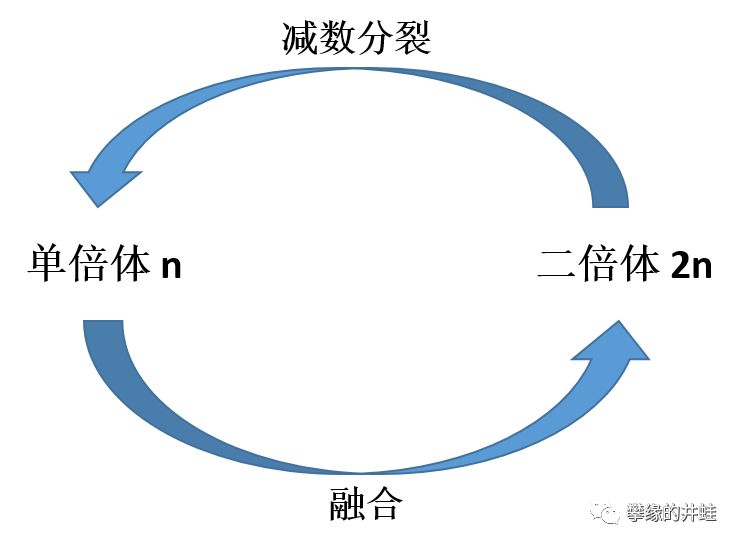
对于所有进行有性生殖的生物来说,它们的生命周期中,一定存在这样一个循环:

融合前的细胞称为单倍体(Haploid),用n表示。融合后,细胞所含的遗传物质增倍,称为二倍体(Diploid),用2n表示。
我们最熟知的n是精子(雄配子)和卵细胞(雌配子)。二者融合成2n的受精卵(合子),受精卵分裂,发育而成的生命体,全部由2n细胞构成,称为二倍体世代,或者孢子体(Sporophyte)世代,为便于描述,后文统称为2n。
此外,有些生物会产生不需要融合(受精)就可以发育成新个体的生殖细胞——孢子。孢子是n,由孢子分裂发育成的生命体全部由n细胞构成,称为单倍体世代,或者配子体(Gametophyte)世代,为便于描述,后文统称为n。
在许多生命的生活史中,n世代和2n世代交替出现,这就是世代交替(Metagenesis)。
然而我们日常看到的绝大多数多细胞生物,包括维管植物的植株,高等真菌的子实体和几乎所有动物,都是它们的2n世代。这些生物不约而同地选择让融合后的2n细胞分裂增殖,分化出不同的形态和功能,发育成具有复杂结构的生命体。而n世代则被不同程度地简化,在极端情况下仅仅作为雌雄配子而存在,沦为2n的附属。
为什么绝大多数多细胞生物选择强化2n?这可能和成对的同源染色体在遗传信息的存储和表达上具有优势有关,本回不做讨论。
自然选择会把微弱的优势不断累积放大。生存竞争越是激烈,这种优胜劣汰越是明显。在弱肉强食的动物界,既不产生孢子,也极少出现能够自由生活的n个体(当然也有例外,以后再详述)。

动物界中的n个体绝大多数由未受精的卵细胞发育而来。最有代表性的比如蜜蜂群中的雄蜂(Drone,上图中)。如果把蜂群看做一个超级生命体,那雄蜂不过是这个超级生命体的生殖器官的一部分,只负责产生精子,同样无法独立生活。图片来源自网络。
而生存压力相对和缓的生产者,比如褐藻,红藻,绿色植物,n大多可以吸收营养,成长发育,与2n组成世代交替。
最典型的世代交替出现在绿色植物的生命周期中。
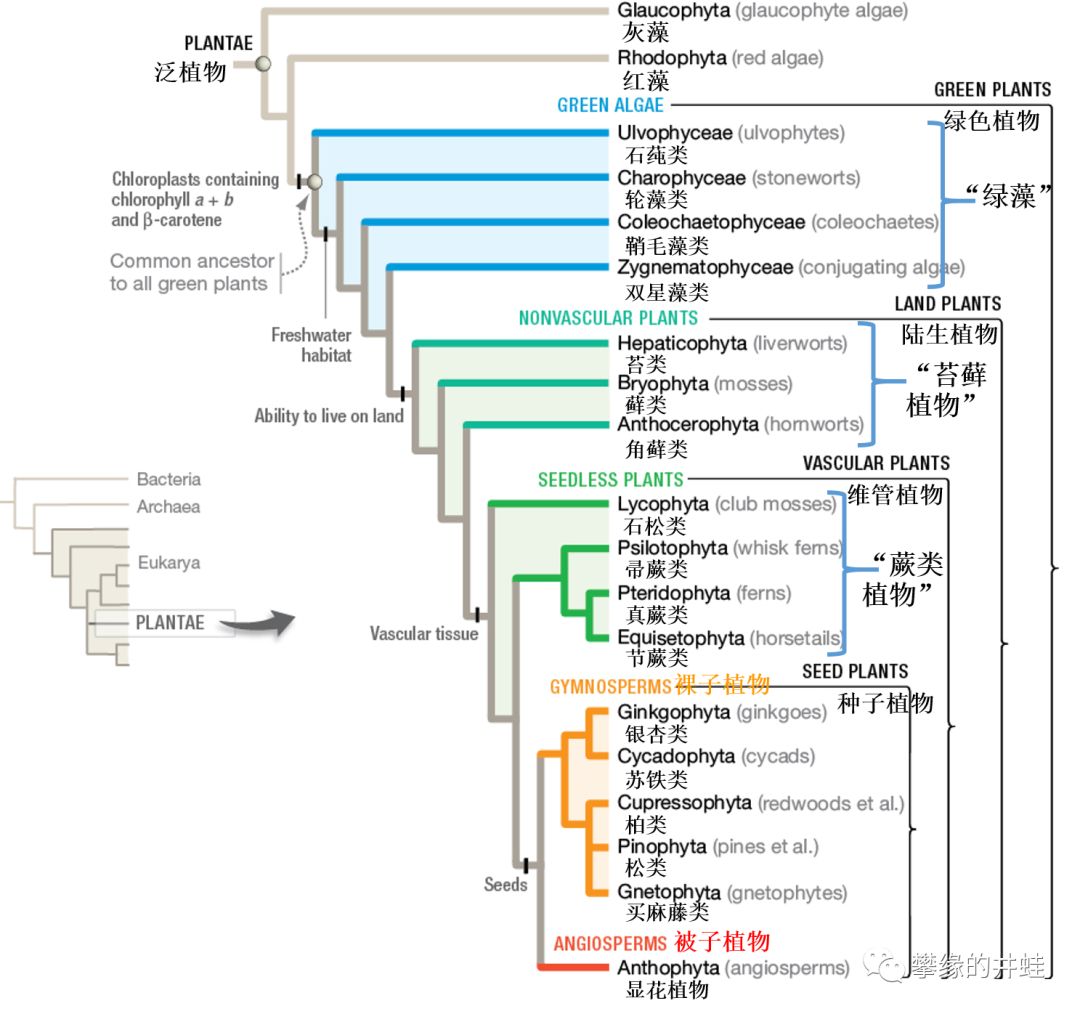
“绿色植物”是“泛植物”下最复杂,最多样,也最成功的一个演化支,包括传统意义上的绿藻,苔藓植物,蕨类植物,裸子植物和被子植物。图片来源自网络。
在一些多细胞绿藻(比如石莼)中,由孢子发育成的n藻体和由合子发育成的2n藻体外观完全相同。这是一种原始的同态(Isomorphic)现象。

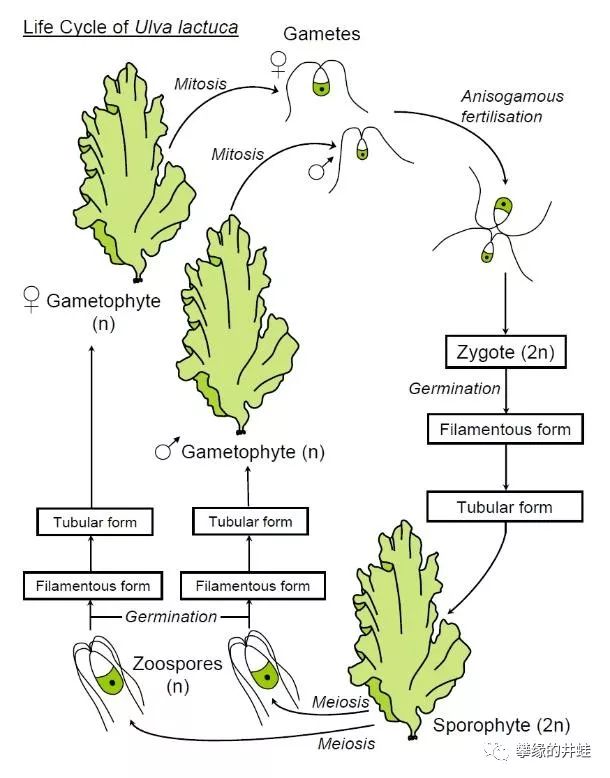
石莼(Ulva)的外形和生活史。不用显微镜观察,你根本无法判断盘子里的这棵海白菜到底是孢子发育成的n,还是合子发育成的2n。图片来源自网络。
就像动物演化出雌性和雄性(雌雄两态),幼体和成体(比如完全变态),营养个体和生殖个体(比如真社会性昆虫)的差异一样,在更高等的绿色植物中,n世代和2n世代也发生差异化,在生命周期中分工协作来提高生存的几率。这种差异化无非两个选择:强化n,或者强化2n。不同的绿色植物类群对这两条演化道路进行了尝试,而且它们的后裔都幸存到了今天。
水生的轮藻和半陆生的苔藓植物采用了强化n的策略。尤其是轮藻,把2n简化到了极致。
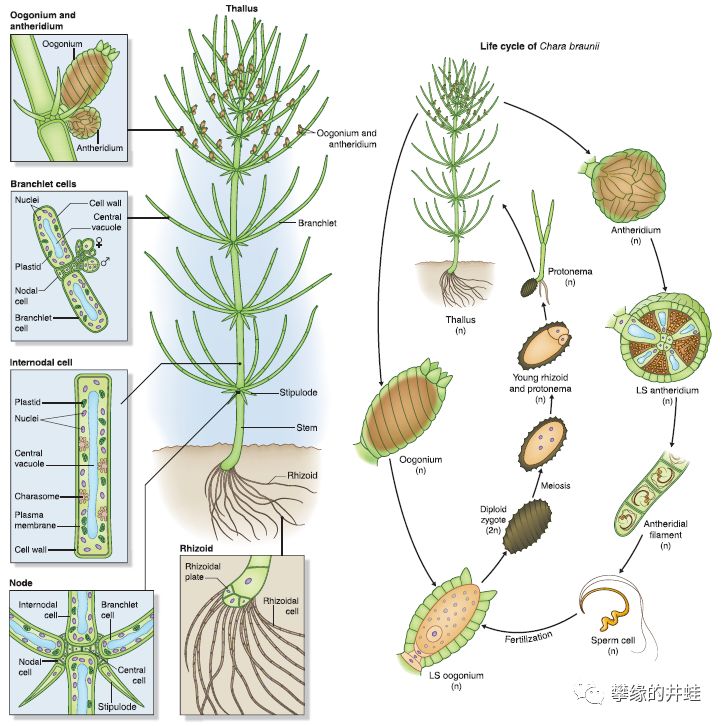
布氏轮藻Chara braunii的形态结构和生活史。注意在它的生命循环中,几乎所有的阶段都是n。只有精子和卵细胞结合后,短暂地形成一个2n细胞:合子。合子的第一次分裂就是减数分裂,重新恢复成n,发育成整株植物。图片来源自[1]。
“苔藓植物”并不是单系群,而是没有演化出维管组织的陆生植物的总称。现存的苔藓植物包括苔类,藓类和角苔类三个分支。它们的合子会分裂发育成一个结构简单的2n世代,2n世代没有根茎叶的分化,不能吸收水分和盐类,也无法进行光合作用。由n(就是我们平常看到的毛绒绒软绵绵的苔藓植株)提供营养生长,可以看作是一个附属的繁殖器官。
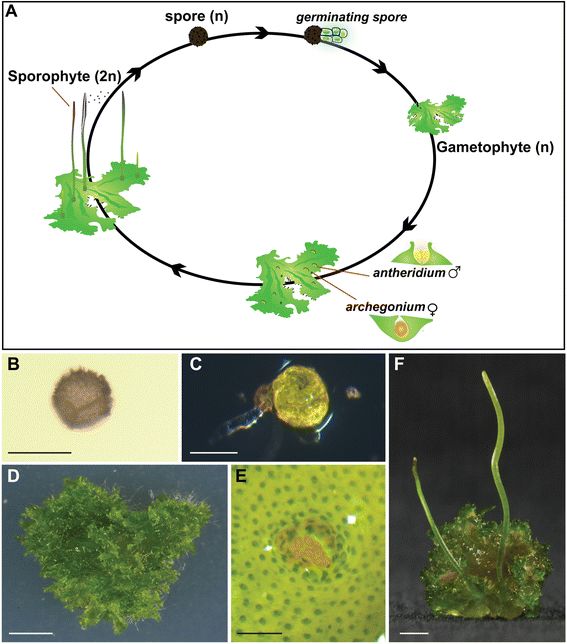
一种角苔Anthoceros agrestis的生活史(A)和外形结构。(B)孢子;(C)萌发中的孢子;(D)配子体(n世代),(E)配子体上的精子囊。F图里那两根细长的棒棒就是它的孢子体(2n世代),寄生在配子体上,唯一的功能就是生产和播撒孢子。图片来源自[2]。
所有的维管植物都采用了强化2n的策略。它们的演化程度更高,也更加成功,这也证明了2n的优势。关于蕨类植物的世代交替,可以参见第一百三十八回 种子。它们的n世代和2n世代都可以独立生活,但是2n更健硕,更复杂,生存时间更长,成为生命循环中的主体。
种子的出现,更加强化了2n的优势地位。
种子的前身是被称为胚珠(Ovule)的微小结构。胚珠的外层是植物母体细心包裹的珠被(2n),中心是由大孢子分裂成的卵细胞和其他几个细胞,这就是种子植物简化到极致的n世代(雌性配子体)。
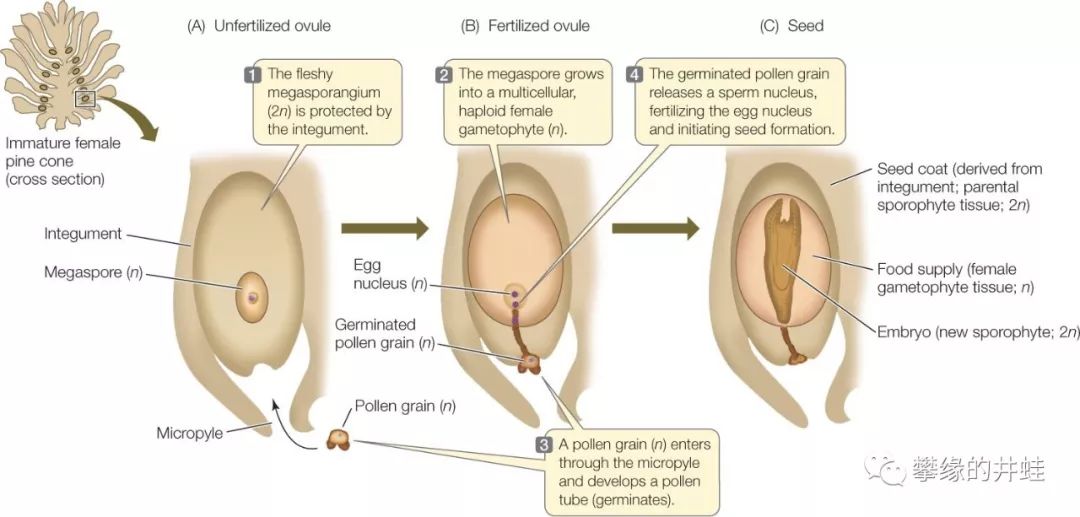
现代松柏类胚珠发育成种子的过程。当花粉中的精子(n)通过花粉管穿过入珠孔(Micropyle),和稳坐珠心(Nucellus)的卵细胞(n)结合,新一代2n世代即宣告诞生,合子(2n)分裂发育成胚(Embryo),也就是一棵完整的植物幼苗;胚珠内的其他n细胞发育成胚乳(Endosperm),储存萌发所需要的营养;2n的珠被发育成种皮,把种子精心封装打包,投送到大自然的怀抱。打个不恰当的比方:种子(2n世代)并不是植物(2n世代)的“子女”,而是“孙子女”。它们中间隔着一个隐藏在胚珠里的n世代。图片来源自网络。
目前发现的最早的胚珠出现在泥盆纪,它们被称为前裸子植物(Progymnospermophyta)。在植物的演化史上,种子很可能不止出现了一次。既然不同的植物类群能够分别独立演化出异型孢子(参见第一百八十八回绿之巨人传4:孢穗),那再进一步,把小孢子变成花粉,把大孢子包裹起来形成胚珠,应该也不是不可能吧。
这也许可以解释为什么许多化石和我们今天看到的植物是那样不同。
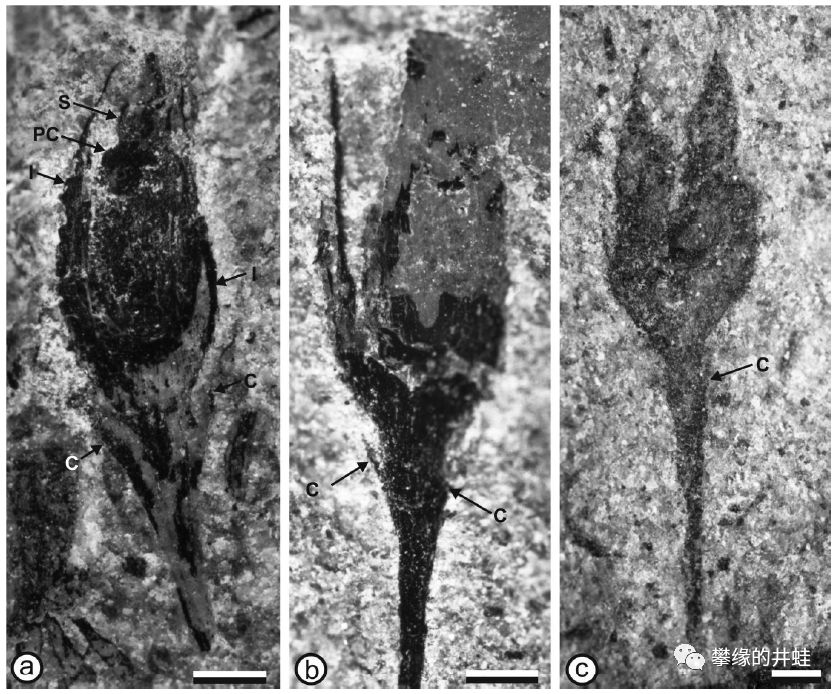
一种泥盆纪种子植物Pseudosporogonites quadrapartitus的胚珠(准确地说是包裹着胚珠的孕育器)化石。它看起来像一朵小花,但其实内部结构完全不同。图片来源自[3],标尺长度1毫米。
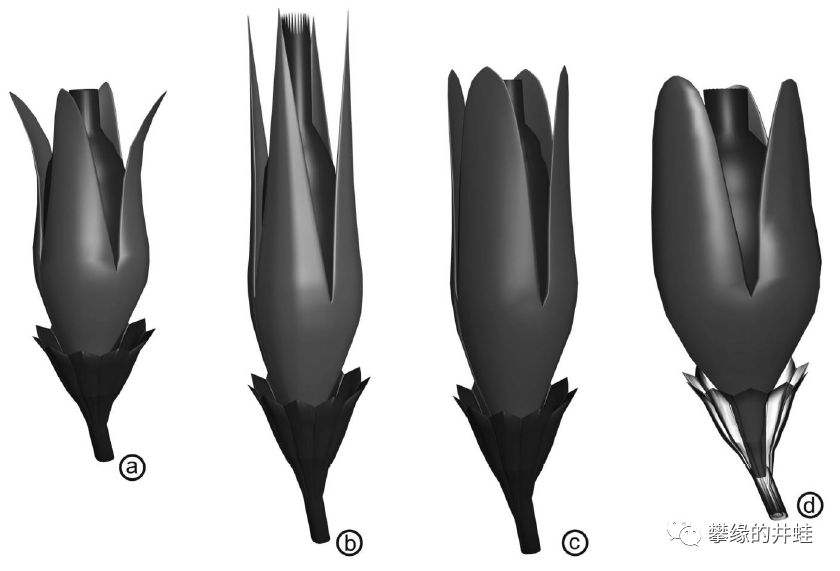
在欧洲和北美都有许多发现了许多Pseudosporogonites属植物化石。上图是P. quadrapartitus(c)和它的几种近亲(a:P. hallei;b:P. bertrandii;d:P. avonensis)孕育器的复原图。图片来源自[3]。
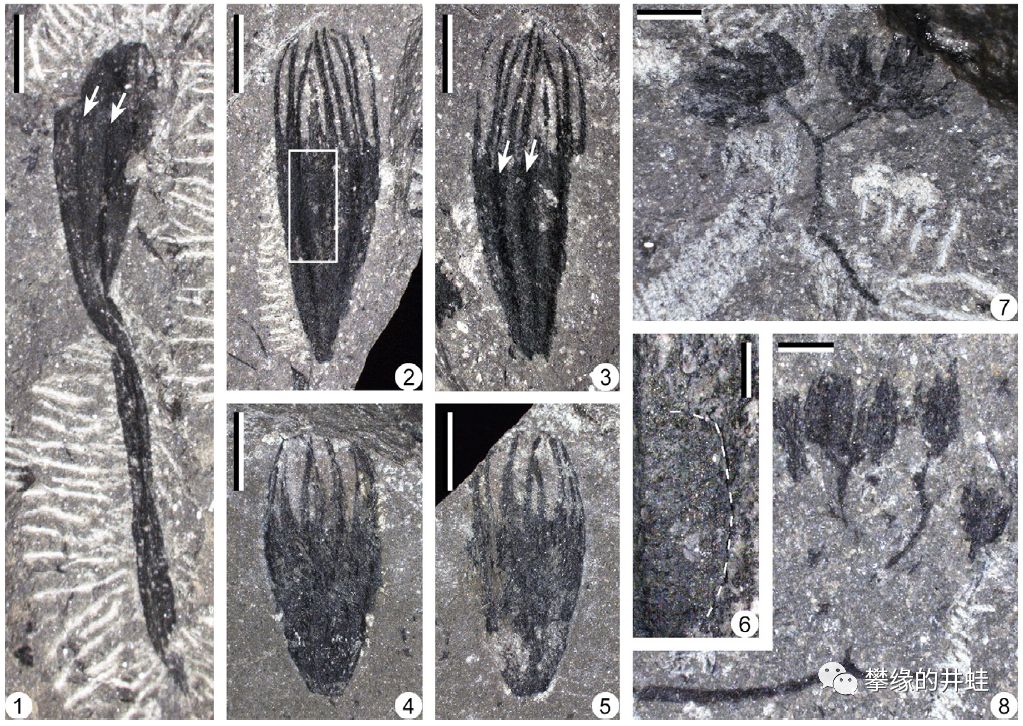
浙江发现的泥盆纪种子植物Calycosperma qii化石。图1到6是包裹胚珠的孕育器(Fertile organ),图7,8是产生花粉的花粉器(Pollen organ)。图片来源自[4]。
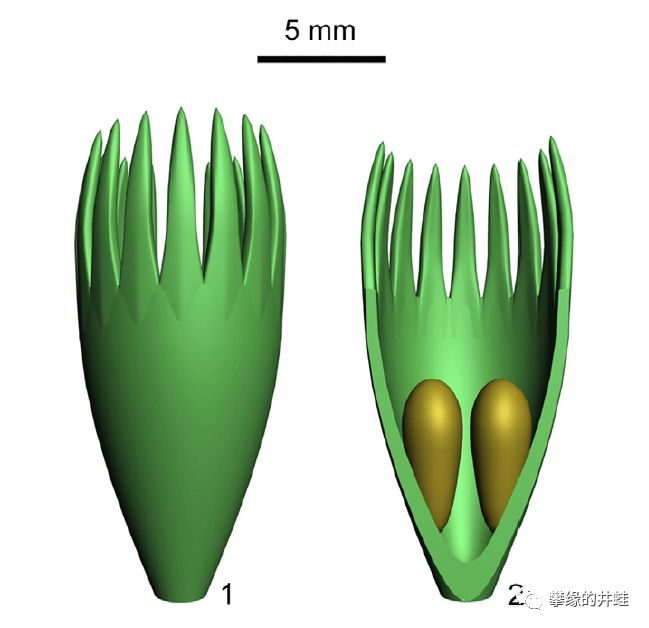
Calycosperma qii孕育器的复原图和剖面结构。每个孕育器里并排生长着两个胚珠。许多泥盆纪种子植物的胚珠外都包裹着这种像萼片一样的结构,可能用来从风中拦截过滤同类的花粉。图片来源自[4]。
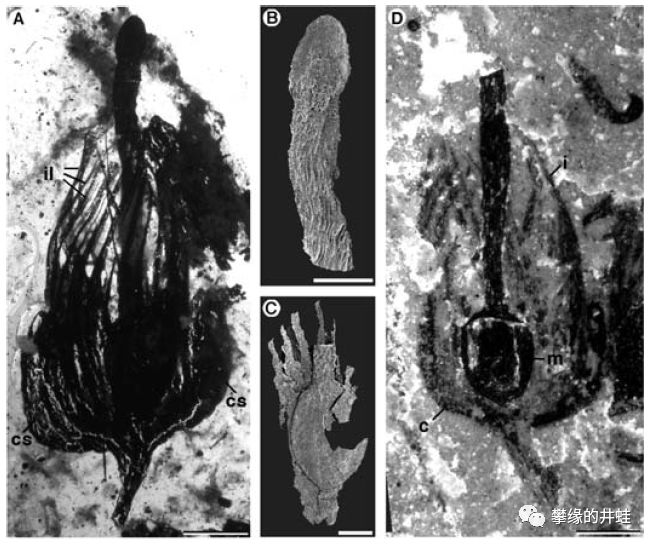
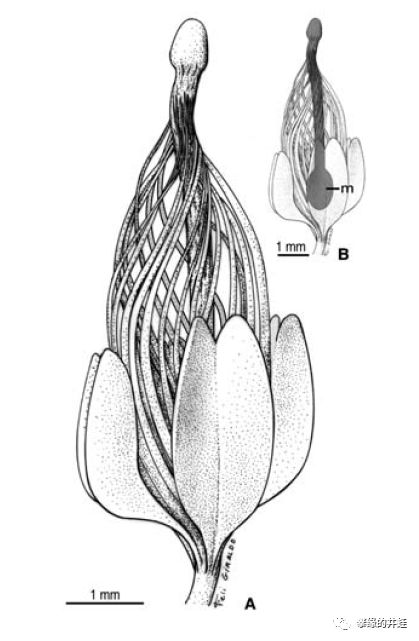
比利时发现的泥盆纪种子植物Runcaria heinzelinii孕育器化石和复原图。中心的m是胚珠的位置,上面顶着一个不太好描述形状的器官,周围丝网状的结构应该也是截留花粉用的。图片来源自[5]。
到了石炭纪和二叠纪,越来越多的种子植物出现。其中一些被认为是现代松柏类(Coniferophyta),苏铁类(Cycadophyta)和银杏类(Ginkgophyta)的祖先。但更多的化石种身世就不那么明朗了,比如乌拉尔山发现的各种似花植物(参见第二百十一回似花还似非花)。它们都被收入裸子植物(Gymnosperm)这个大筐。之所以称为“裸子”,是因为更加高级的植物演化出更复杂,更华丽的组织来包裹胚珠(这是中生代的故事),相比之下,这些早期种子植物的胚珠简陋单薄,近乎“裸露”。
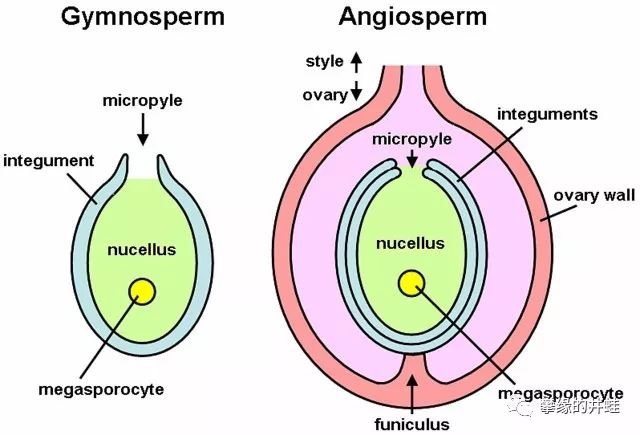
左:裸子植物的胚珠结构。右:被子植物(Angiosperm,就是各种显花植物)的胚珠外面,还有一层被称为子房(Ovary)的套装。胚珠发育成种子,子房发育成果实。图片来源自网络。
俄罗斯彼尔姆地区发现的早二叠世种子植物Astrogaussia imbricata种子的复原图。它所在沃尔茨杉目(Vojnovskyales)在古生代晚期颇为繁盛,但它们的分类地位并不明确。图片来源自[6],标尺长度1厘米。


意大利多洛米蒂山脉(Dolomites)发现的晚二叠世种子植物Pernerina pasubi化石和复原图。它具有扇形的叶片(像银杏),细长的繁殖叶上长有两排种子。也是植物演化史上一个身份不明的神秘过客。图片来源自[7]。
井蛙最感兴趣的,是种子蕨类(Pteridospermophyta)。许多年以前,井蛙自己还是个孩子的时候,第一次在书上看到“种子”和“蕨”,这两个似乎毫无关系的词语组合在一起。在收集本回的素材时,当时的那种新奇与兴奋,又一次在心底苏醒了。
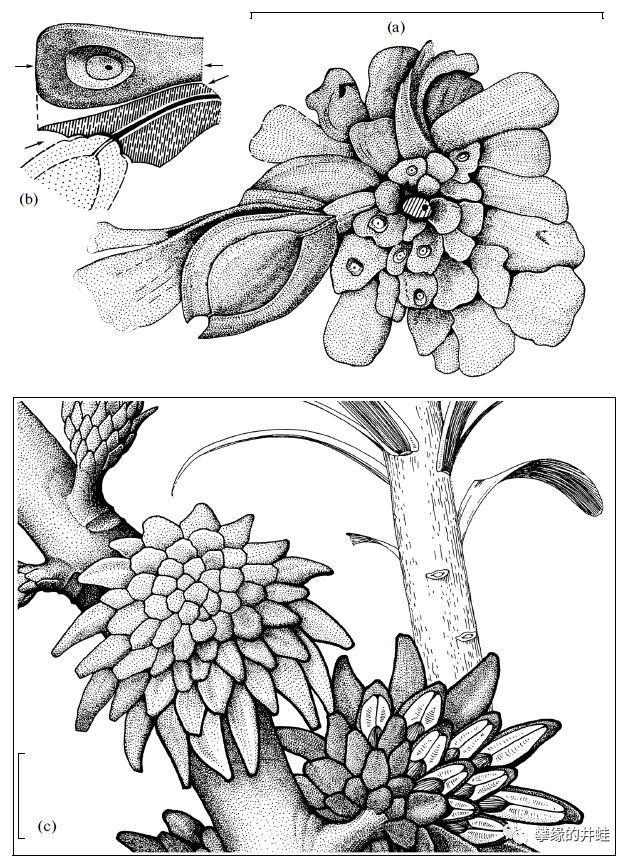
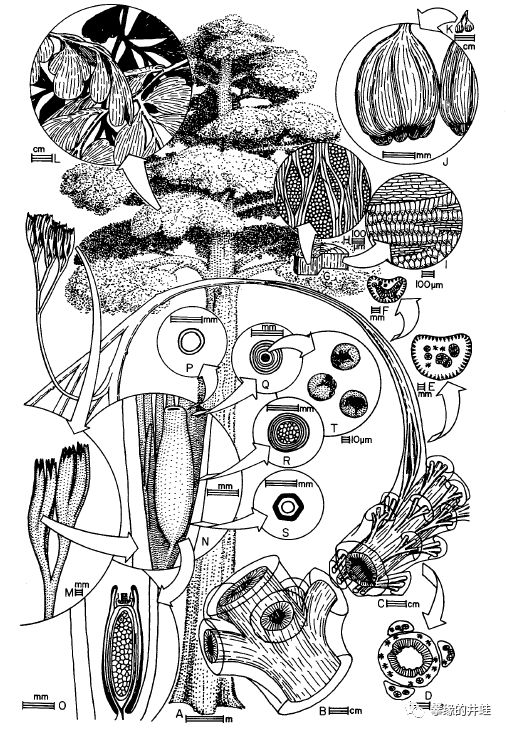
种子蕨是石炭纪到二叠纪期间重要的成煤植物,留下的大量化石资料,包括各种乔木,灌木,藤蔓和草本。井蛙从文献[8]扒来了一些精美的复原图,以飨读者。大型植物几乎不可能留下完整的化石,都是分散的根系,枝干,叶片,还有花粉器,孕育器和种子。研究者们为了复原这些植物的全貌,花费了无数心血。
Stamnostoma huttonense,生长在早石炭世的苏格兰。
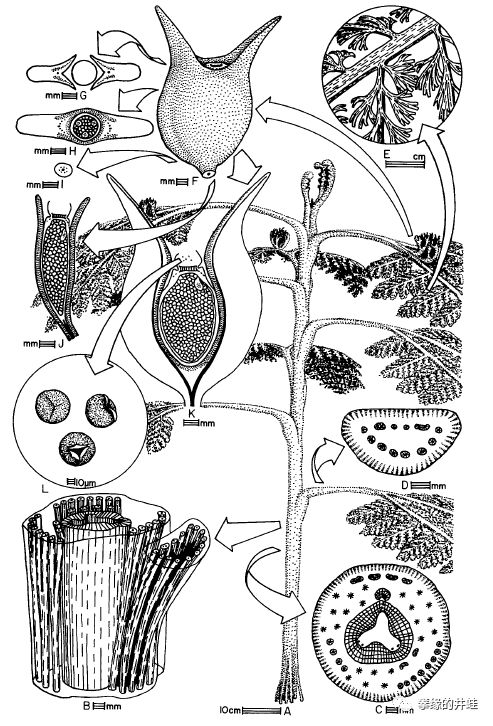
Lyraspermascotica,生长在早石炭世的苏格兰。
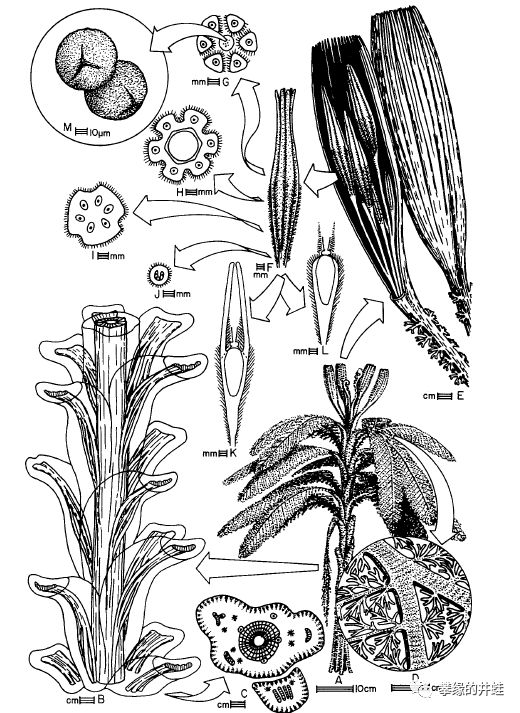
Calathospermumfimbriatum,生长在早石炭世的苏格兰。

Lagenostoma(瓶籽属)lomaxii,生长在早石炭世的英格兰。

Pachytesta(小厚壳籽属)illinoensis。晚石炭世,伊利诺伊州。

Callospermarion(靓籽属)pusillum。晚石炭世,伊利诺伊州。

Dictyopteridium sporiferum,晚二叠世,澳大利亚昆士兰。
一般来说,根据胚珠和种子化石定名的种子蕨,叫“某某籽”;根据叶片化石定名的种子蕨,叫“某某羊齿”。其中自然难免出现同一种植物被重复命名的情况。比如下面的例子:

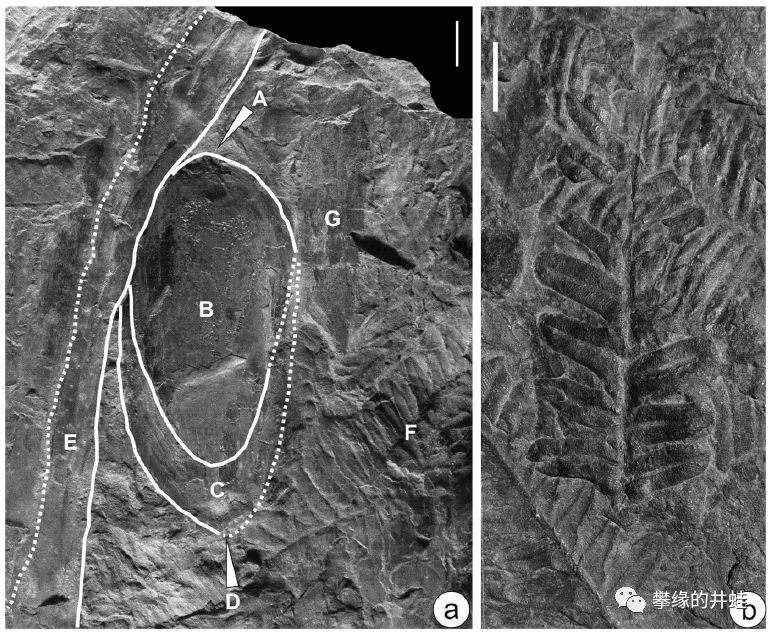
这块晚石炭世化石上保留了一段植物的印痕,在蕨类羽叶簇拥的枝干上,挂着一枚完好的种子(白色箭头指示,见下面的大图)。发现地:罗马尼亚Reşiţa Basin。有趣的是,这种植物的种子和叶片之前被分别定名为小厚壳籽Pachytesta incrassata和座延羊齿Alethopteris zeilleri。现在人们知道了:它们红莲白藕青荷叶,本来是一家。图片来源自[9]。
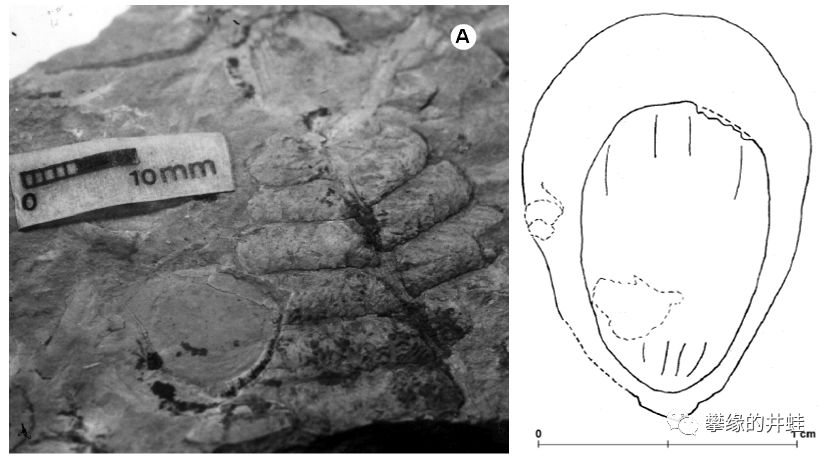
这块古巴出土的早二叠世化石清晰地显示了一枚种子长在一片蕨类植物的叶片上。之前这种叶片被命名为Delnorteacf. D.abbottiae,属于大羽羊齿科(Gigantpoterideae)。而种子被归入高特籽属(Cordaicarpus)。又有两块碎片被成功拼接到了一起。图片来源自[10]。
不同的种子蕨,种子外形和生长位置也是千奇百怪,各不相同。比如在加拿大Sydney Coalfield,就发现了这些化石:

一种三棱籽属(Trigonocarpus),胚珠和叶子上的小羽片并排生长。图片来源自[11]。
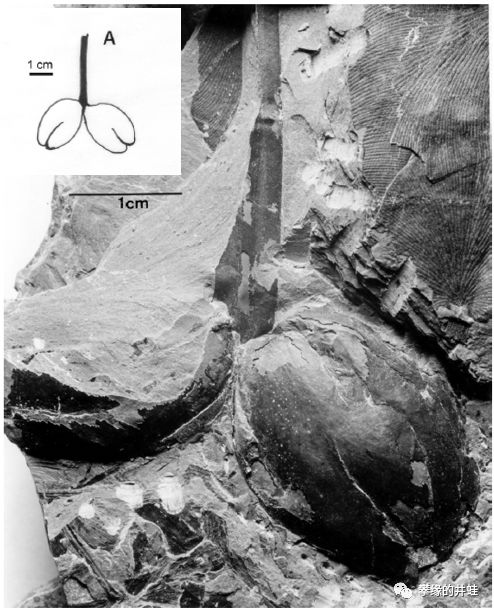
另一种三棱籽属,两个胚珠一起长在长柄顶端。图片来源自[11]。
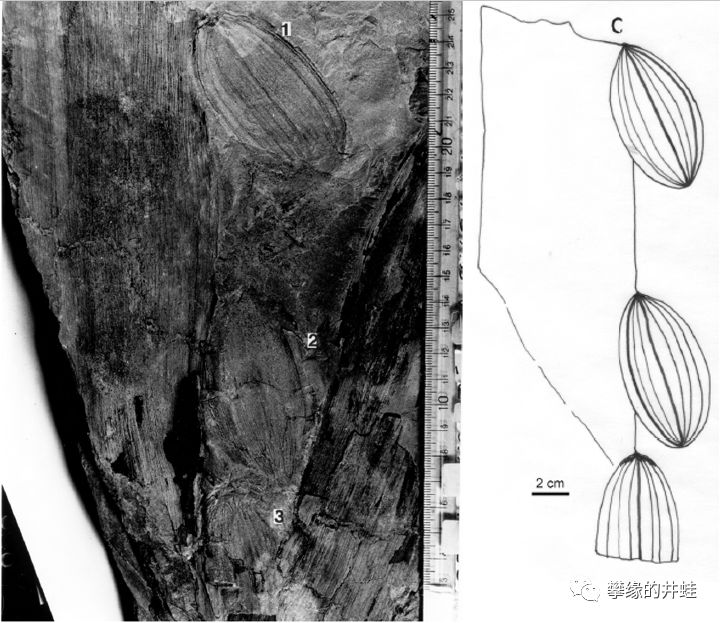
一种小厚壳籽属Pachytesta incrassata,胚珠成排长在叶片边缘。图片来源自[11]。
这些卵形的附属物也许看上去像是某种奇怪的包块或者虫瘿(参见第一百八十七回六足传奇7:蝗食蜩饮),但化石剖面结构证明,它们是不折不扣的胚珠和种子。
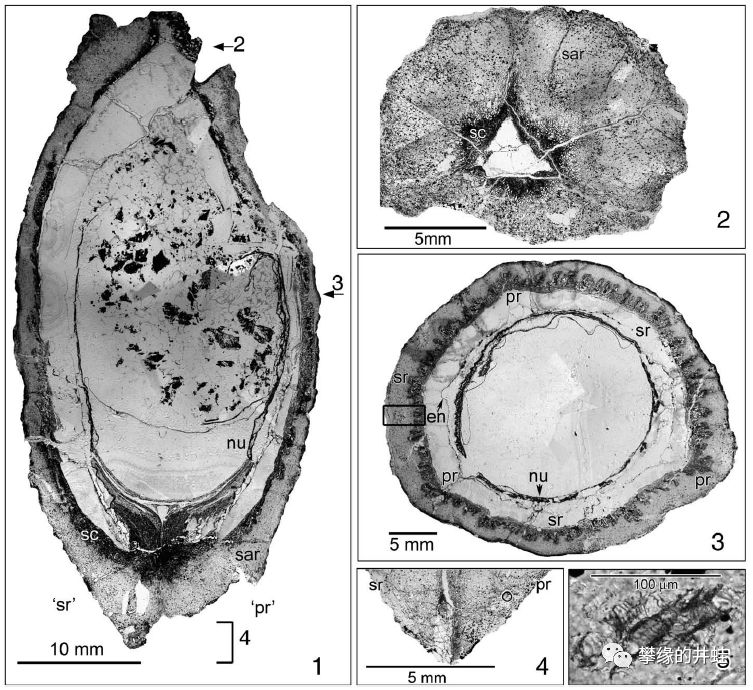
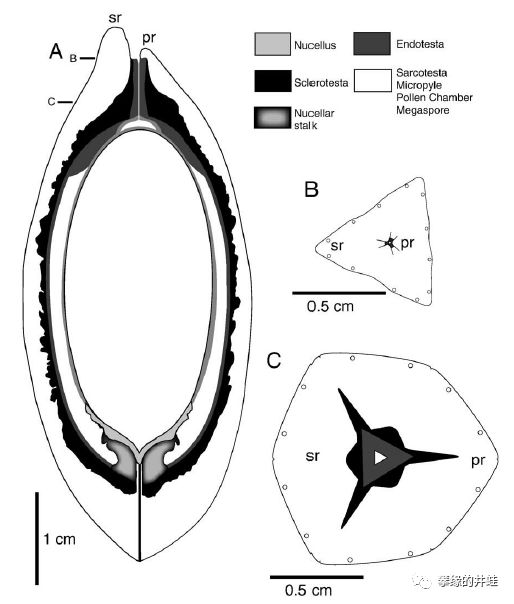
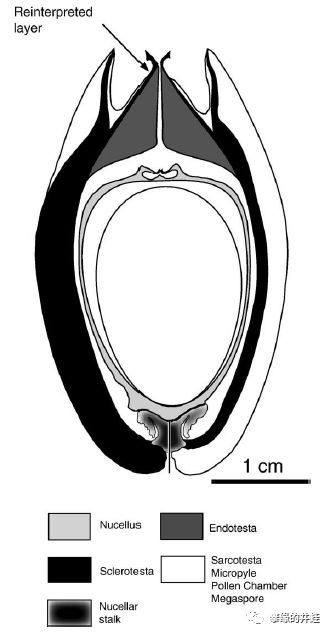
晚石炭世小厚壳籽属Pachytesta crenulata胚珠化石剖面和复原图。发现地:爱荷华州Shuler and UrbandaleMines。可以清晰地看到包裹胚珠的珠被(Integument,会发育成种皮),花粉停留的花粉室(Pollen chamber),让花粉管通过,传递精子的通道,等等。注意它的中心没有发育成型的幼苗(胚),表明这是一个尚未受精的胚珠(n),还没有发育成种子(2n)。图片来源自[12]。
另一种晚石炭世小厚壳籽属Pachytesta illinoensis(上面有它的完整复原图)的胚珠结构。比Pachytesta crenulata结构更复杂,生存压力也在迫使种子蕨不断对胚珠和种子进行改进。图片来源自[12]。
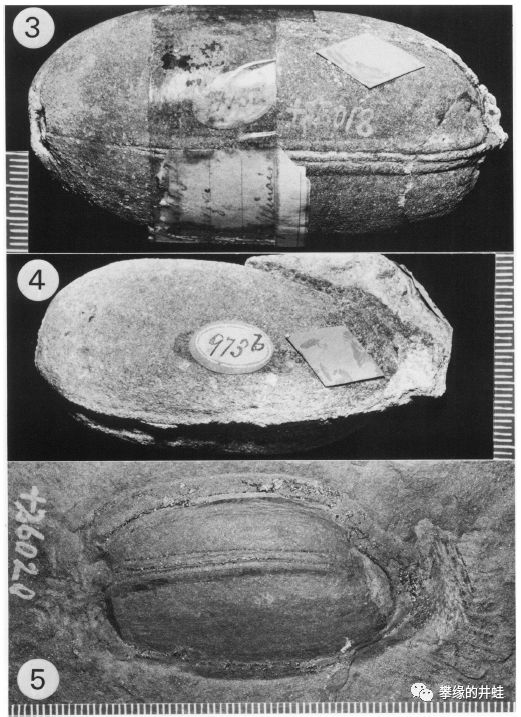
三棱籽属Trigonocarpus leeanus(晚石炭世,伊利诺伊州)的种子化石。种皮厚实坚固,有明显的筋肋,有点像今天的坚果。当时应该还没有四足动物能攻破它的盔甲,所以这样的结构可能用来保存水分,抵御炎热干旱低温等不良环境,防止昆虫蛀蚀,等等。图片来源自[13]。
种子蕨是一个非常多样,也非常成功的植物类群。它们一直生存到白垩纪晚期(甚至有疑似新生代种子蕨化石发现)才最终退场。期间出现了无数种类。本文只能挂一漏万,蜻蜓点水地稍加介绍。最后,再放上一种奇特的种子蕨,来结束本回。
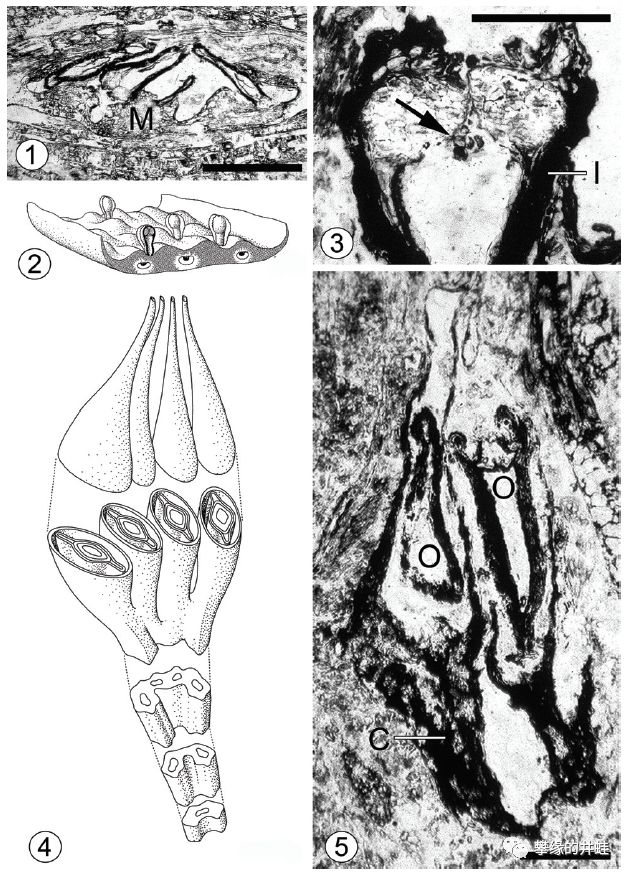
南极洲比尔德莫尔冰川(Beardmore Glacier)发现的晚二叠世舌羊齿类Glossopteris胚珠化石剖面和复原图。在它的大孢子叶上并排生长这4个胚珠。授粉后将发育成四胞胎种子。身处冈瓦纳古陆的南端,幸运地躲过了西伯利亚暗色岩事件的直接冲击,它和它的近亲们将把种子蕨家族延续到恐龙时代。图片来源自[14],标尺长度1毫米。
地球名片
生物分类:泛植物界-绿色植物-陆生植物-维管植物-前裸子植物
存在时间:泥盆纪早期?至石炭纪早期
现存种类:无
化石种类:不明
生活环境:陆地
代表特征:兼具真蕨类,种子蕨和裸子植物(松柏类,苏铁类)的特征,分类地位不明,可能是种子植物的先驱
代表种类:戟枝木类,原髓木类,古羊齿类
[PSSP] R. Luthringer, A. Cormier, S. Ahmed,et al., Sexual dimorphism in the brown algae. Perspectives in Phycology, Vol. 1(2014), Issue 1, p. 11–25
[1] Tomoaki Nishiyama, Hidetoshi Sakayama,Jan de Vries,et al., The Chara Genome: Secondary Complexity and Implicationsfor Plant Terrestrialization. Cell 174, 448–464, July 12, 2018 449
[2] Péter Szövényi, Eftychios Frangedakis,Mariana Ricca, et al., Establishment of Anthoceros agrestis as a model speciesfor studying the biology of hornworts. BMC Plant Biology (2015), 15:98, DOI10.1186/s12870-015-0481-x
[3] Cyrille Prestianni, Jason Hilton, WaltCressler, Were All Devonian Seeds Cupulate? A Reinvestigation ofPseudosporogonites hallei , Xenotheca bertrandii, and Aglosperma spp. InternationalJournal of Plant Sciences, Vol. 174, No. 5 (June 2013), pp. 832-851
[4] Le Liu, De-MingWang, Mei-Cen Meng, etal., A newseed plantwithmulti-ovulate cupules fromthe Upper Devonian of SouthChina. Review of Palaeobotany and Palynology, 249 (2018), 80–86
[5] P. Gerrienne, B. Meyer-Berthaud, M.Fairon-Demaret, et al., Runcaria, a Middle Devonian Seed Plant Precursor.SCIENCE, 29 OCTOBER 2004, VOL 306, DOI: 10.1126/science.1102491
[6] S. V. Naugolnykh, Fossil Flora andStratigraphy of the Terrigenous Kungurian Beds (Lower Permian) of the Basin ofthe Barda River (Urals, Perm Krai). Stratigraphy and Geological Correlation, 2014,Vol. 22, No. 7, pp. 680–707
[7] Michael Wachtler, Pernerina pasubi nov.gen. n. sp. an enigmatic plant of unknown botanical affinity from the LatePermian Wuchiapingian (Lopingian) of the Dolomites (Northern Italy).Dolomythos, 2015
[8] Gregory J. Retallack, David L Dilcher,Reconstructions of Selected Seed Ferns. Annals of the Missouri BotanicalGarden, 75: 1010-1057, 1988, DOI: 10.2307/2399379
[9] Christopher J. Cleal, Mihai E. Popa, Position of Ovule Attachment in aCarboniferous Medullosalean Pteridosperm. Botanica Pacifica. A journal of plantscience and conservation, 2015. 4(2): 127–130, DOI: 10.17581/bp.2015.04209
[10] Ricardi, F., O. Rösler, O. Odreman,Tafoflora de Delnortea (Gigantopterideae) de Loma de San Juan (FormaciónPalmarito, NO de Venezuela) y sus relaciones paleofitogeográficas en elArtinskiano (Neopaleozoico). PLANTULA 2(1-2): 73-86
[11] Erwin L. Zodrow, Note on differentkinds of attachments in trigonocarpalean (Medullosales) ovules from thePennsylvanian Sydney Coalfield, Canada. Atlantic Geology 40, 197–206 (2004)
[12] Anne Raymond, Rachel McCarty,Pachytesta crenulata, a new medullosan ovule from the Pennsylvanian of Iowa.Review of Palaeobotany and Palynology, 156 (2009), 283–306,doi:10.1016/j.revpalbo.2009.03.007
[13] Robert A. Gastaldo and Lawrence C.Matten, TRIGONOCARPUS LEEANUS, A NEW SPECIES FROM THE MIDDLE PENNSYLVANIAN OFSOUTHERN ILLINOIS. Amer. J. Bot. 65(8): 882-890, 1978
[14] Taylor, E.L., T.N. Taylor, and P.E.Ryberg (2007), Ovule-bearing organs of the glossopterid seed ferns from theLate Permian of the Beardmore Glacier region, Antarctica, in Antarctica: AKeystone in a Changing World – Online Proceedings of the 10th ISAES, edited byA.K. Cooper and C.R. Raymond et al., USGS Open-File Report 2007-1047, ShortResearch Paper 082, 4p.; doi:10.3133/of2007-1047.srp082