
种子植物在二叠纪蓬勃发展。寒冷干燥的风掠过泛古大陆,从无数千奇百怪的花粉器(Pollen organ)上带走微小的花粉颗粒,播撒到同样千奇百怪的孕育器(Fertile organ)上。两方的遗传物质在胚珠中融合,发育成更加千奇百怪的种子,再生长为愈发千奇百怪的植株。地球上第一次百“花”齐放。


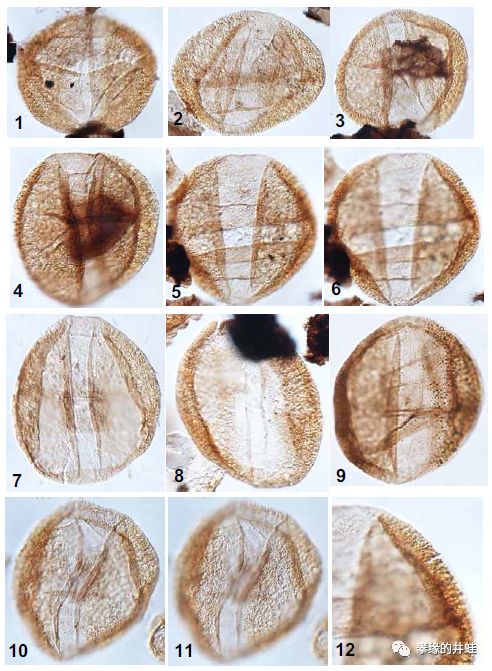
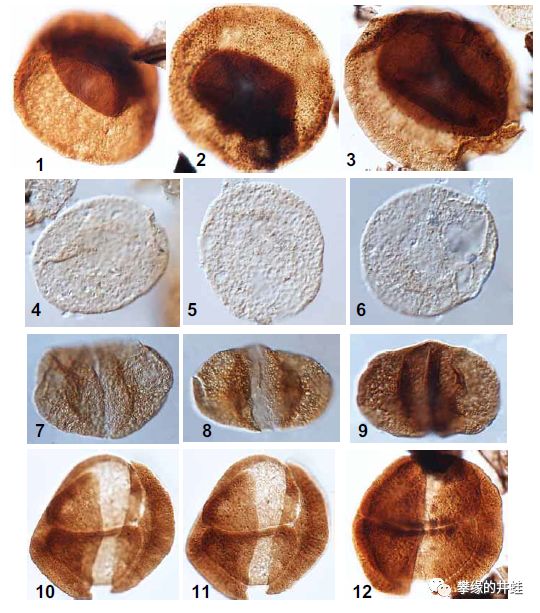
阿曼和沙特发现的早二叠世花粉颗粒化石。和同时期的孢子相比,花粉颗粒的外壳更薄,形态也更加轻巧,许多花粉还装备了“小翅膀”,更适合随风飘荡。图片来源自[1]。
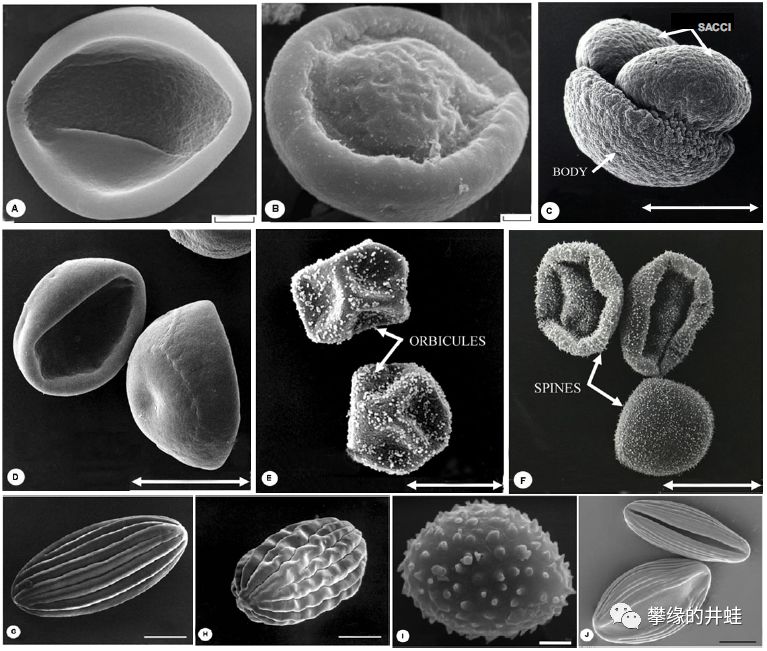
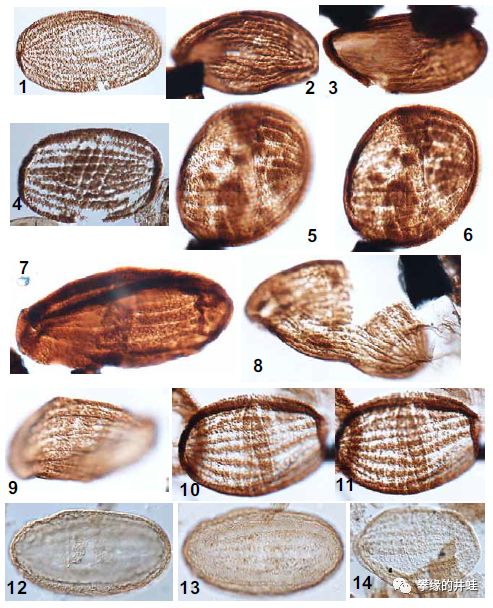
一些现代裸子植物的花粉颗粒照片。(A)叉叶苏铁Cycas micholitzii;(B)银杏Ginkgo biloba;(C)Pinus ponderosa;(D)美国黄松Pseudotsuga menziesii;(E)黄桧Chamaecyparis nootkatnesis;(F)西部铁杉Tsuga heterophylla;(G-H)美洲麻黄Ephedra americana;(I)买麻藤Gnetum macrostachyum;(J)百岁兰Welwitschia mirabilis。和3亿年前的花粉前辈对比一下,有些似乎有那么一点点相似。图片来源自[2]。
在世界各地的化石发现地找到了各种各样的二叠纪种子植物,大多被扔到了“种子蕨”这个筐里——这也表示没有找到它们和现存植物的明确的亲缘关系。但也有一些化石,研究者根据胚珠,孢子叶和维管束的形态,认定为现代裸子植物——苏铁类(Cycads),银杏(Ginkgo,注意这个词不是复数)和松柏类(Conifers)的早期种类。
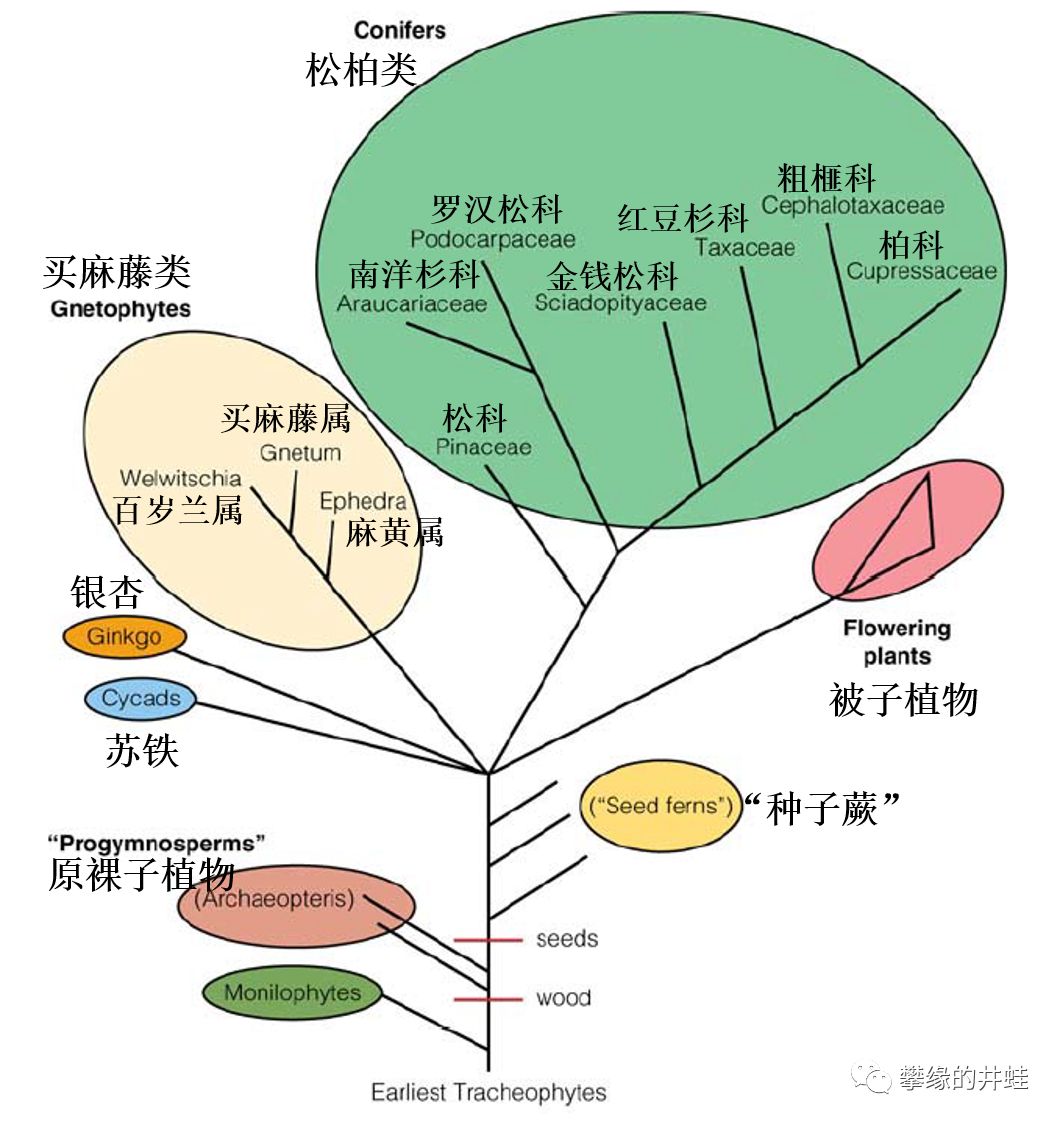
种子植物演化关系极简图。要理清各个类群的演化关系非常困难,目前比较确定的,是现存的被子植物和裸子植物构成了一个单系群。也许历史上“种子”曾经独立出现过几次,但现存的所有种子植物都起源自同一个祖先。“裸子植物”是本图的重点。广义上的“裸子植物”包括各种分类不明的种子蕨,以及被子植物的祖先。狭义的“裸子植物”包括苏铁类,银杏和松柏类三大类群。图中还画上了一个比较冷门的买麻藤类(Gnetophytes),从前被当做现存裸子植物的第四大类群,不过新的基因检测结果显示它们其实是松柏类中的一个小分支。图片来源自[3]。
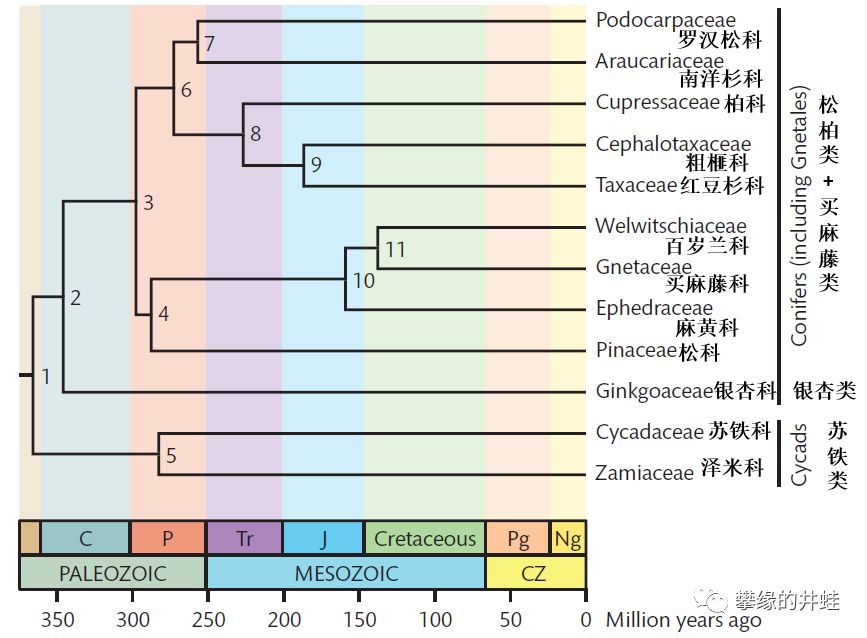
根据基因测序结果绘制的现代裸子植物亲缘关系和演化时间。大约在泥盆纪末期,苏铁类的祖先从种子植物基干中分化出来。随后,银杏类和松柏类的祖先在石炭纪早期分道扬镳。到二叠纪,三大类群都已经演化成型。买麻藤类是松柏类在中生代演化出的一支。这些结果也和目前发现的化石资料相吻合。图片来源自[4]。
苏铁门(Cycadophyta)有大约300个现存种。我们最熟悉的要数琉球苏铁(Cycas revoluta)。因为树形古朴美观,四季常青,种植简单,分株方便,寿命长而且病虫害少等等优点,作为景观树被大量栽种。不过,琉球苏铁在北方属于养死不容易,养好更不容易的哪一类。所以才会有成语“铁树开花”,用来形容事情稀少或者困难。井蛙是到宁波后,才看到苏铁开“花”的样子,才知道大的苏铁树每年都会开花结籽。


上图:苏铁属植物的“雄花”(小孢子叶球)。下图:苏铁属植物的“雌花”(大孢子叶球)和发育中的种子。图片来源自网络。

a:台东苏铁(Cycas taitungensis)附着在大孢子叶上的种子,你看它又大又圆。b,c:华南苏铁(Cycas rumphii)附着在大孢子叶上的胚珠。图片来源自[5]。

苏铁(Cycas sexseminifera)的大孢子叶球(a),生长在大孢子叶上的胚珠(b,c),和发育成熟的种子(d)。图片来源自[6],标尺长度1厘米。井蛙在宁波也曾经扒过琉球苏铁的“雌花”,看到杏那么大那么红的种子长在金黄的“叶子”上,当时就想:这不就是传说中的“种子蕨”吗?
作为最靠近裸子植物基干的分支,苏铁类是现存植物中唯一大孢子叶(Megasporophylls)保持着叶片形态的类群。从这个角度上看,它们确实是现存植物中最接近“种子蕨”的。事实上,有研究者认为苏铁类和晚石炭世的种子蕨类籽羊齿属(Spermopteris)关系密切。除了“长在叶子上的种子”,苏铁类还保持着其他一些 “原始”的特征,比如聚生在树顶的羽叶,还有布满树干表面的叶柄痕迹。这让它们看上去(只是看上去)非常接近蕨类植物和裸子植物的过渡形态。
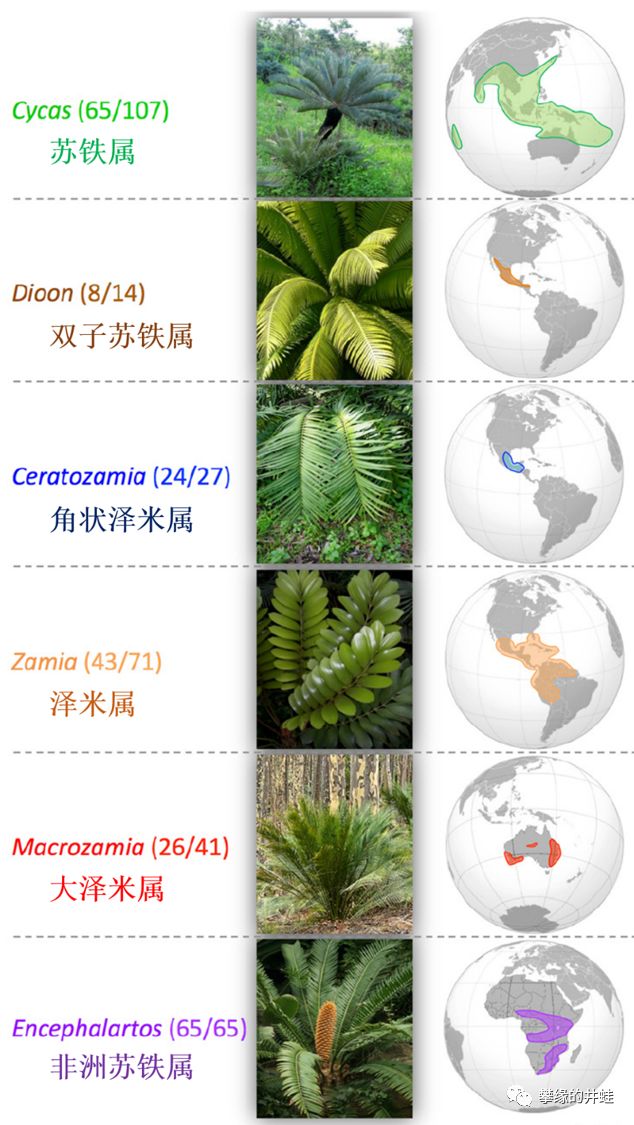
现存苏铁的主要属,和野生种群的分布区域。这些古老植物的孑遗星星点点地散布在热带和亚热带地区。图片来源自[7]。
从化石记录来看,苏铁类似乎最早出现在温热潮湿的华夏植物区(Cathaysia Floral Province,参见第二百三十二回绿之巨人传6:孤岛)。在我国发现了世界上最早的苏铁类化石,为研究苏铁类的起源和早期演化提供了珍贵的证据。
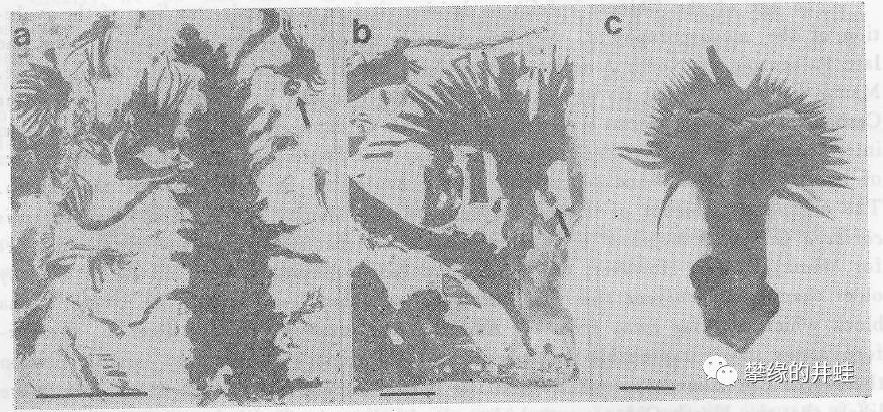
山西发现的目前已知生存年代最早的苏铁类化石:早二叠世晚期的中国始苏铁(Primocycas chinensis)大孢子叶化石(a,b)。箭头指示了上面附着的胚珠。c是现代暹罗苏铁(Cycas siamensis)的大孢子叶和胚珠。图片来源自[8],标尺长度1厘米。文献是1989年的,图片质量比较感人。井蛙查到同一化石点还发现了另一种苏铁类:古生铁花Cycadostrobilus paleozoicus, 化石保存为小孢子叶球,文献[9],可惜没有找到图片。

另一种山西发现的早二叠世苏铁类Crossozamia chinensis(中文名应该是中国什么泽米?)大孢子叶和胚珠(a,标尺长度1厘米)化石。这张照片清楚多了,和现代苏铁的大孢子叶对比一下,结构非常相似。图片来源自[5]。
到二叠纪晚期,苏铁类已经在泛古陆广泛分布,形貌也日趋多样。



意大利北部多洛米蒂山脉Dolomites 发现的晚石炭世苏铁类Nilssonia brandtii化石和复原图。它的大孢子叶球和种子形态很接近现代的泽米科(Zamiaceae)苏铁。很可能在二叠纪,现代苏铁类的主要类群就都已经出现了。图片来源自[10]。


鳞秕泽米(Zamia furfuracea,原产墨西哥)的外形(上图)。它的大孢子叶球和种子聚成一根玉米棒似的圆柱(下图)。图片来源自网络。
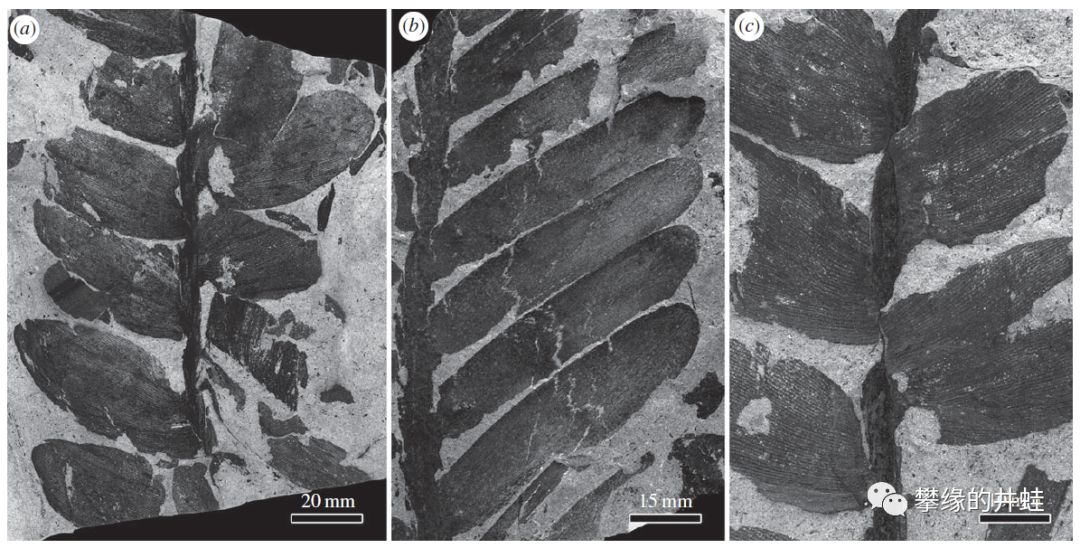
云南发现的晚二叠世苏铁类Plagiozamites oblongifolius叶片化石。它的叶片形态和气孔结构和现代泽米科苏铁非常接近。图片来源自[11]。

山西发现的早二叠世苏铁类Pseudoctenis samchokense叶片化石。图片来源自[12],标尺长度1厘米。研究者曾经认为它属于苏铁门的另一个主要分支:本内苏铁目(Bennettitales或Cycadeoideales),另一个分支是苏铁目(Cycadales),包括各种苏铁和泽米。不过最新的研究显示它更接近苏铁目。本内苏铁目在三叠纪到侏罗纪极尽繁盛,等到中生代的章回中再详细介绍。
苏铁门虽然没落了,总算还有300来种生存到今天。差不多同时出现的银杏门(Ginkgophyta)就没有这么幸运了。现存的就只剩下一根独苗苗:银杏纲(Ginkgoopsida)银杏目(Ginkgoales)银杏科(Ginkgoaceae)银杏属(Ginkgo)的银杏(Ginkgo biloba)。

作为银杏门唯一的现存物种,银杏虽然被尊为国宝活化石,虽然被大量引种到世界各地,但仍然掩盖不住那份孤寂与凄凉。小扇子一样的叶片是银杏最显著的特征,到秋天就会变成美丽的金黄色(b),经常被夹入书页,多年后不经意间飘落出来,唤醒一段沉睡的记忆。(a)展示了生长在叶簇上的胚珠,就是那些绿色的小球。银杏门也在中生代兴盛一时。(c)是河南发现的三种侏罗纪银杏类复原图。左为义马银杏(Ginkgo yimaensis),右上为义马果(Yimaia recurva),右下为河南卡尔肯果(Karkenia henanensis)。它们展示了银杏门下曾经多么“树才济济”。图片来源自[13]。
银杏门的始祖可能和石炭纪出现的两歧叶属(Dichophyllum)有关。比较确定的早期银杏门植物包括二叠纪银杏类有楔拜拉属(Sphenobaiera)和毛状叶属(Trichopitys)。从外形上看,它们和现代银杏差别还是挺大的。

法国洛代夫盆地(Lodève Basin)发现的二叠纪植物化石,疑似一种两歧叶Dichophyllum moorei。银杏的祖先很可能就长成这种样子的。图片来源自[14],标尺长度1厘米。


多洛米蒂山脉发现的两块晚二叠世楔拜拉属银杏类化石(学名未定)。可以看到它们的种子(胚珠)也是长在叶子上的。看来大家都经历过“种子蕨”的演化阶段呢。图片来源自[15],标尺长度1厘米。
现代银杏已经脱胎换骨。它的大孢子叶退化,和其他的裸子植物相比,银杏的种子更像是被子植物的浆果。也许是殊途同归吧,从中生代开始,各种古代银杏也演化出肥厚多汁的外种皮(注意不是果皮果肉)来吸引动物食用,顺便(真的是顺着便便)撒播种子,大大提高了生存能力。然而福兮祸所伏,也许正是这种高度同质化的竞争策略,让银杏遇到真正的被子植物时一败涂地。当然,这是后话了。

银杏被大量种植在许多大学,社区和街道。每到深秋,大量成熟的白果(其实是种子)掉落地面,腐烂发酵,空气中弥漫一种特殊的酸涩味道。现在,一种特殊的无毛灵长类承担起为仅存的银杏传播种子的任务。图片来源自网络。
说个趣闻吧:研究者曾经在日本发现一株银杏,胚珠下长出了大孢子叶,发现者把它命名为“O-ha-tsuki(日语应该写作“お葉つき”吧?)”。看上去就像是种子长在了叶片上,结构真有点像二叠纪的楔拜拉属银杏。

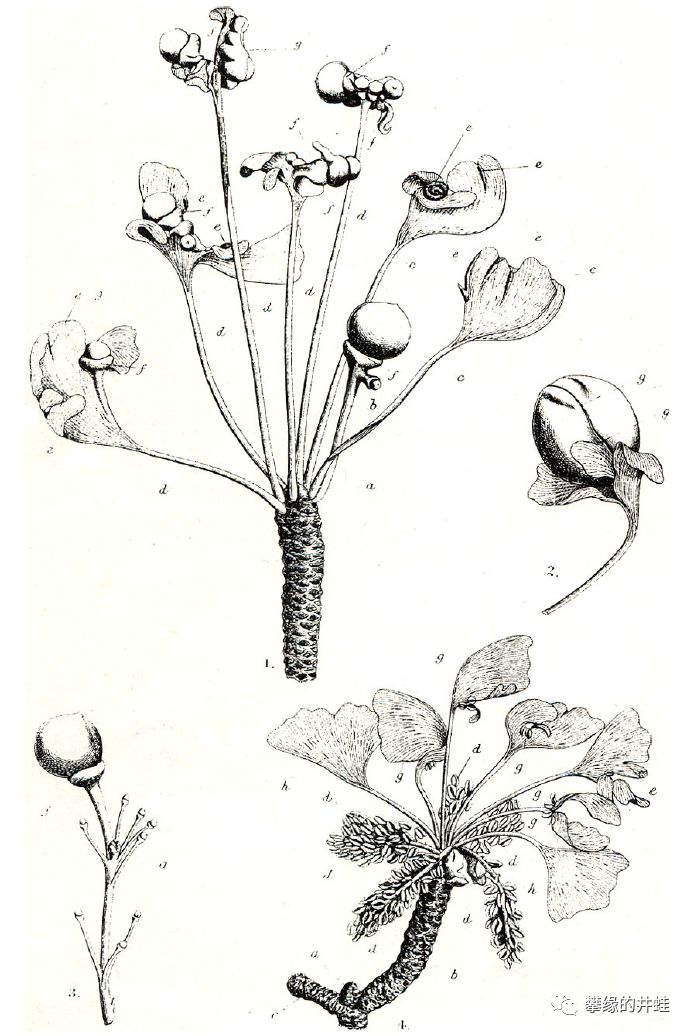
“お葉つき?”的标本和记录画图。这可能是一例极其少见的返祖现象,某个或某几个沉睡了亿万年的基因意外地苏醒表达,把银杏演化史上的某个瞬间稍微展现了一下。图片来源自[15]。
再说说松柏门。今天大约有630种现存松柏杉桧榧(买麻藤类并入松柏门后,还要再加上大约80种)分布在世界各地,是当之无愧的裸子植物杠把子。基本上,一提起“裸子植物”,大家首先想到的肯定是各种松柏。和苏铁门和银杏门相比,松柏门出现时间要稍微晚一点。早期的松柏类——科达木(Cordaites)在石炭纪已经和木本石松,节蕨类,真蕨类和种子蕨一起,构筑起连绵无尽的泥炭森林。石炭纪末雨林崩溃事件后,科达木逐渐衰退,新的松柏类群兴起,二叠纪的松柏类多样性已经非常可观了。

多洛米蒂山脉出土的晚石炭世松柏类化石。a:Ortiseia leonardii;b:Ortiseia visscheri;c-d:Ortiseia jonkeri;e,k:Pseudovoltzia liebeana;f:Dolomitia cittertiae;g:Quadrocladus Schweitzer;h:未定种松柏类孢子叶球;i:疑似Leptostrobus;j:松柏类树干;l:木炭;m-o:树干细胞微结构。图片来源自[16]。
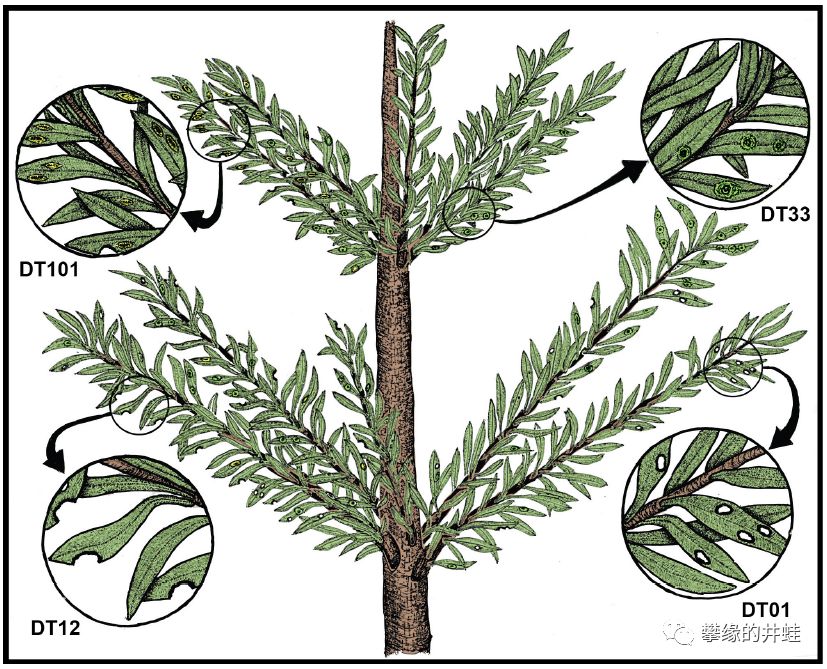
Pseudovoltzia liebeana复原图。这种早期松柏类的化石资料非常丰富,包括叶片上被多种昆虫啮咬和寄生的痕迹(参见第一百八十七回六足传奇7:蝗食蜩饮)。可以看到,早期松柏类的叶片都比较普通,并不是我们熟悉的针状或者鳞片状。图片来源自[16]。

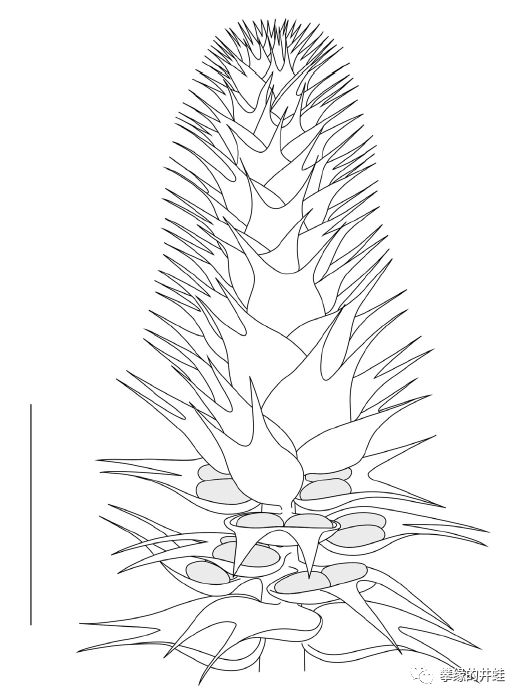
印尼苏门答腊岛发现的早二叠世松柏类Tobleria bicuspis的大孢子叶球化石和复原图。属于伏脂杉目(Voltziales),大约介于科达木和现代松柏类之间。这个大孢子叶球已经很有点像今天的松塔了。但每片孢子叶上生有两枚种子。图片来源自[17],复原图标尺长度1厘米。
二叠纪的松柏类还不能凌霜傲雪四季常青(当然合成一块的泛古大陆也不大可能有四季分明的气候)。在冈瓦纳古陆的南端高纬度地区确实有大片寒带林,但其中的主要树种却是一类种子蕨——舌羊齿类(Glossopterids)。

南极洲Lamping Peak 发现的二叠纪树桩和根系化石。这些树桩基本保存完好,就像乌达煤田的“植物庞贝城”(参见第二百三十回 绿之巨人传6:孤岛)一样,为研究者提供了植物种类,密度,生长情况等等宝贵信息。图片来源自[18]。

二叠纪Lamping Peak寒带林复原图。在那个时代,无论合弓纲还是蜥形纲,应该都无法在如此寒冷的地区生存。栖息在林中的大概只有一些昆虫之类的节肢动物。寒冷,深邃,静谧。图片来源自[18]。
西伯利亚的超级火山爆发宣告了一个时代的结束。和地球上所有的生灵一样,陆生植物也经历了一轮残酷的大洗牌。当这场史无前例的灭绝事件的尘埃落定,苏铁类,银杏类和松柏类迎来了自己的时代。千姿百态的苏铁和银杏为各种食草巨兽提供了充足的食物,供养起中生代庞大富饶的陆地生态系统。松柏类则取代种子蕨,在高海拔高纬度的地区迎击霜雪,傲然屹立。直到今天,针叶松柏仍然是寒带造林的绝对主力。
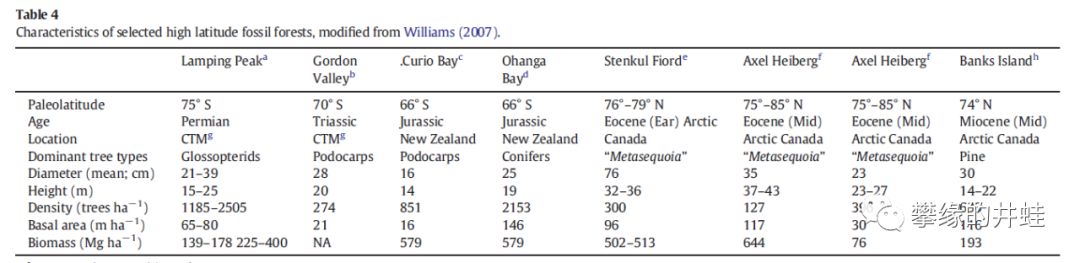
高纬度地区化石森林统计表(摘自[18])。在主要树种(Dominant tree types)一行,只有二叠纪是舌羊齿(一类种子蕨)。三叠纪是罗汉松(Podocarps),侏罗纪是罗汉松和针叶树(Conifers),新生代是水杉(Metasequoia)和松(Pine),都是松柏类的常绿树。
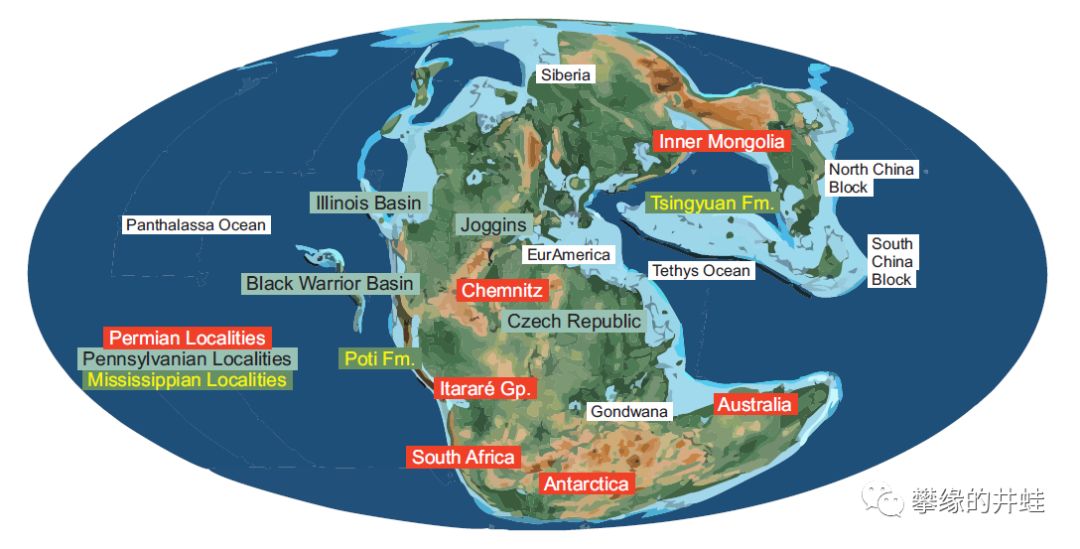
至此,以石松类,节蕨类,真蕨类和种子蕨为代表的石炭-二叠纪成煤植物的故事结束了。我们再来看一看晚古生代的世界地图,和主要植物化石发现地。在新的时代,无论海陆分布还是植物形态都有很大不同。中生代陆生植物的故事依然精彩,将在其他系列中继续讲述。图片来源自[19]。
地球名片
生物分类:泛植物界-绿色植物-陆生植物-维管植物-裸子植物
存在时间:泥盆纪早期?至现代
现存种类:约1010种(松柏门约630种,苏铁门约300种,银杏门1种,买麻藤类约80种)
化石种类:未找到完整统计
生活环境:陆地
代表特征:胚珠直接生长在大孢子叶上,有胚珠无子房,有种子无果实
代表种类:种子蕨,苏铁类,银杏类,松柏类
[1] MICHAEL H. STEPHENSON, SPORES ANDPOLLEN FROM THE MIDDLE AND UPPER GHARIF MEMBERS, PERMIAN, OMAN. British Geological Survey
[2] Danilo D. Fernando, Christina R. Quinn,Eric D. Brenner, et al., Male Gametophyte Development and Evolution in ExtantGymnosperms. International Journal of Plant Developmental Biology, 2010
[3] Davis, Gymnosperms, University of California
[4] Susanne Renner, Gymnosperms. TheTimetree of Life, Oxford Biology
[5] Miao Y, Liu ZJ, Wang M, Wang X (2017)Fossil and Living Cycads Say "No More Megasporophylls". J MorpholAnat 1: 107
[6] Xin Wang, Bin Luo, Mechanical Pressure,Not Genes, Makes Ovulate Parts Leaf-Like in Cycas. American Journal of PlantSciences, Vol.4 No.12A(2013), Article ID:41272, DOI:10.4236/ajps.2013.412A1008
[7] FabienL Condamine, Nathalie S Nagalingum, Charles R Marshall, et al., Origin and diversificationof living cycads: a cautionary tale on the impact of the branching processprior in Bayesian molecular dating. BMC Evolutionary Biology (2015), 15: 65,DOI 10.1186/s12862-015-0347-8
[8] GAO ZHI-FENG, BARRY A. THOMAS,Occurrence of earliest Cycads in the Permian of China and its bearing on theirevolution. Chinese Science Bulletin, Vol. 34(1989), No. 9
[9] Zhu Jia-nan, Zhang Xiu-sheng, Ma Jie. ANew Genus and Species—Cycadostrobilus paleozoicus Zhu of Cycadaceae from thePermian of China. , 1994, 32(4): 340-344
[10] Michael Wachtler, The cycad Nilssoniabrandtii n. sp. from the Late Permian Wuchiapingian (Lopingian)Gröden-Formation (Dolomites - Northern Italy). Dolomythos, 2015
[11] Feng Z, Lv Y, Guo Y, et al., Leafanatomy of a late Palaeozoic cycad. Biol. Lett. 13(2017): 20170456,http://dx.doi.org/10.1098/rsbl.2017.0456
[12] Pott, C., McLoughlin, S., andLindstrom, A. 2010. Late Palaeozoic foliage from China displays affinities toCycadales rather than to Bennettitales necessitating a re−evaluation of thePalaeozoic Pterophyllum species. Acta Palaeontologica Polonica 55 (1): 157–168
[13] Crane PR. An evolutionary and culturalbiography of ginkgo. Plants, People, Planet, 2019; 1: 32–37
[14] J. Galtier, J. Broutin, Floras fromred beds of the Permian Basin of Lodève (Southern France). Journal of IberianGeology, 34 (1) 2008: 57-72
[15] Thilo C Fischer, Barbara Meller,Evelyn Kustatscher, et al., Permian ginkgophyte fossils from the Dolomitesresemble extant O-ha-tsuki aberrant leaf-like fructifications of Ginkgo bilobaL. BMC Evolutionary Biology 2010, 10:337
[16] Evelyn Kustatscher, Johanna H.A. vanKonijnenburg-van Cittert, Cindy V. Looy, et al., The Lopingian (late Permian)flora from the Bletterbach Gorge in the Dolomites, Northern Italy: a review.Geo. Alp, Vol. 14, 2017
[17] Van Waveren IM (2019) A morphometricanalysis of Tobleria bicuspis, a Voltziales seed cone from the early PermianJambi palaeoflora, Sumatra (Indonesia). PhytoKeys 119: 67–95
[18] Molly F. Miller, Nichole E. Knepprath,David J. Cantrill, et al., Highly productive polar forests from the Permian ofAntarctica. Palaeogeography, Palaeoclimatology, Palaeoecology 441 (2016)292–304
[19] R.A. Gastaldo, Marion Bamford, JohnCalder, et al., UNIT 12: The Coal Farms of the Late Paleozoic.