
从这一回开始,我们把镜头切回龙兽争霸的现场。
在二叠纪,羊膜动物刚刚取得陆地霸权不久,它们的体型远不如中生代和新生代的后辈们那样庞大,外形也没有那么花样百出。许多类群还保持着灰头土脸的“蜥蜴”的样子。
想象一下,井蛙和几个朋友回到二叠纪的泛古大陆。仔细确认附近没有危险后(二叠纪中后期已经出现了能对成年男子团体构成威胁的大型肉食合弓兽,从那时起,穿越者们再也不能无忧无虑地在野外乱逛了),大家一边聊天一边散步观景。突然发现一条“蜥蜴”懒懒地趴在地上。没费什么力气就抓住了它。仔细观察之后,大家认定:这是某种鳞鲵(参见第二百三十一回 不尴不尬),属于“古两栖动物”壳椎类。好家伙,连羊膜动物都不是。
放走了鳞鲵后,没过多久,又发现一条“蜥蜴”。抓来研究之后,认定这是一条蜥代龙(参见第二百二回 百兽图3:鸭子),属于合弓纲中比较原始的盘龙类,和咱们算是远亲。
放走了蜥代龙,没过多久,双发现一条“蜥蜴”。抓来研究之后,认定这是一条杨氏蜥(参见第二百五回 水陆空V1.0),属于蜥形纲中的早期双孔类,算是真正的蜥蜴的远祖了。
放走了杨氏蜥,没过多久,叒发现一条“蜥蜴”。抓来研究之后,认定这是一条大鼻龙(参见第二百四回 蛀牙,断尾巴),属于蜥形纲真爬行动物中一个比较原始的类群。
放走了大鼻龙,没过多久,叕发现一条“蜥蜴”。抓来研究之后,认定这是一条波罗蜥,属于蜥形纲中的副爬行动物。
放走了波罗蜥——哎~别忙,你还没说清楚呢,啥叫“副爬行动物”?还有刚才的“真爬行动物”是怎么回事?
咳咳,副爬行动物(Parareptilia)和真爬行动物(Eureptilia)是蜥形纲下的两个演化支。大体上,真爬行动物包括传统分类学上的“双孔类(Diapsida)”,以及和双孔类近缘的其他类群,囊括了几乎所有你能想到的“爬行动物”:蜥蜴,蛇和沧龙(鳞龙类),鳄类(镶嵌踝类主龙),龟鳖(现在也被证明是双孔类),鱼龙(鱼龙类),蛇颈龙(鳍龙类),翼龙(鸟颈类主龙),恐龙(鸟颈类主龙),还有鸟类(就是恐龙)。怎么看上去好像也没给副爬行动物留下什么啊?确实是这样,因为副爬行动物在二叠纪末损失惨重,少数残余也在三叠纪末全部灭绝,蜥形纲在中生代和新生代的大放异彩和它们完全没有关系。正因如此,这个演化支只好屈居为“副”,总好像低人一头的意思。
但在二叠纪,副爬行动物才是蜥形纲的“正统”和“主流”。虽然同样被合弓纲压制,副爬行动物无论数量,体型大,还是多样性,都比同时期的真爬行动物高出不少,甚至有迹象表明,它们开始染指一些合弓纲和离片椎类占据生态位。

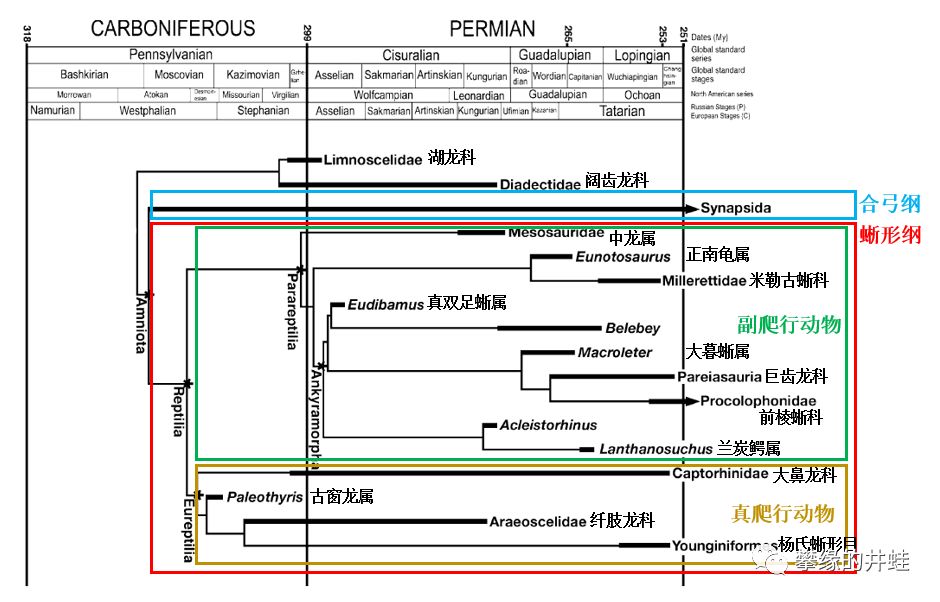
副爬行动物的主要类群和生存时代。这张图中把正南龟属(Eunotosaurus)归入副爬行动物,是比较有争议的。图片来源自[1]。

南非卡鲁盆地发现的正南龟(Eunotosaurus africanus)化石。宽扁的肋骨很容易让人联想到正在成形中的龟壳,不过也有可能只是趋同演化。关于正南龟,一种观点认为它是副爬行动物的早期分支,属于米勒古蜥总科(Millerettidea)中比较特化的一类;另一种观点认为它是龟鳖形类(Testudinata)的早期成员,应该放到真爬行动物那边去。以后写龟鳖类起源和演化的时候还会重点介绍这货。图片来源自[2]。
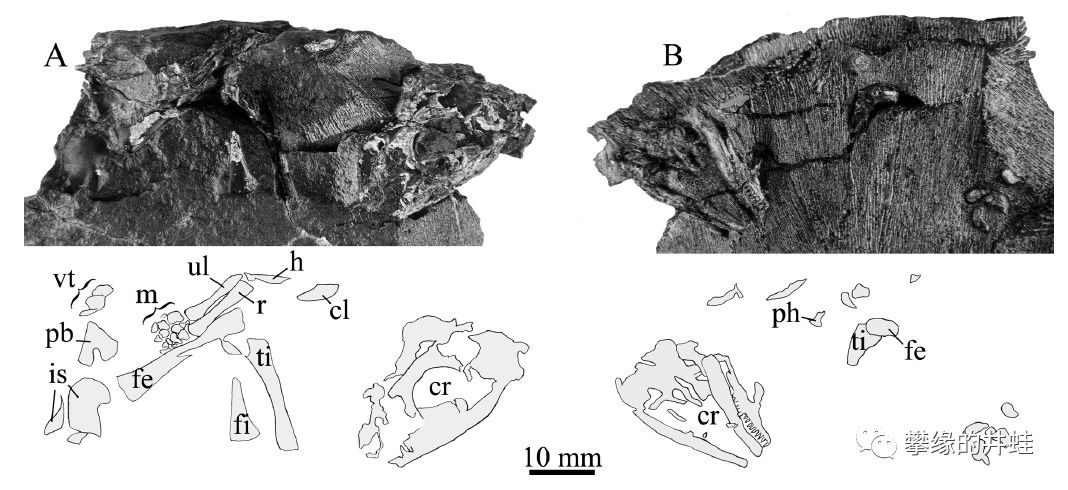
南非发现的晚二叠世米勒古蜥Broomia perplexa化石,保存了破碎的头骨,骨盆和右后肢。它和正南龟生活在同一时期,同一地区,算是久别重逢的老邻居了,但不知道二者沾亲不。图片来源自[3]。
除了包括米勒古蜥类,目前公认的副爬行动物还有波罗蜥类(Bolosauria),中龙类(Mesosauria),前棱蜥类(Procolophonoidea,包括夜守龙Nyctiphruretus),巨齿龙类(有些文献也译作锯齿龙类,Pareiasauria),和兰炭鳄类(Lanthanosuchoidea)等。
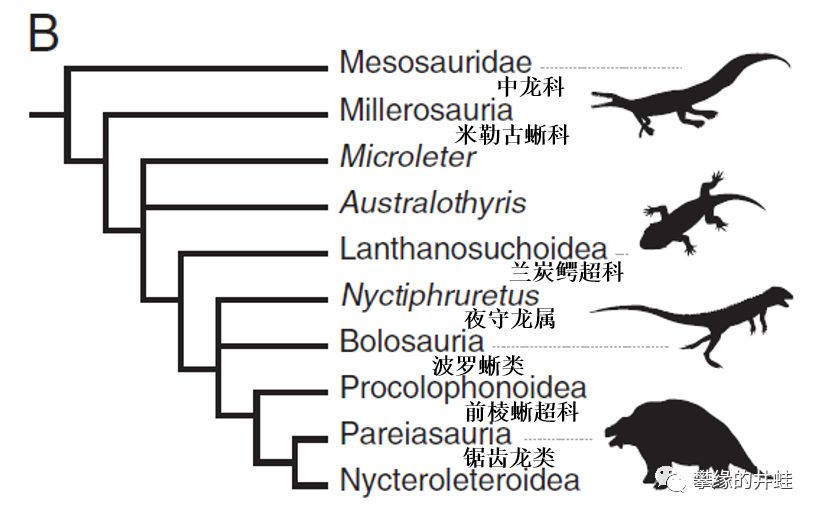
关于副爬行动物演化谱系的另一个观点。右边的剪影从上到下:纳米比亚中龙Mesosaurus tenuidens,沃氏兰炭鳄Lanthanosuchus watsoni,真双足蜥Eudibamus cursoris,盾皮龙Scutosauruskarpinskii。从这也能看出来,二叠纪的副爬行动物家族挺兴旺的,有些身影似曾相识。图片来源自[4]。
从结构上看,二叠纪的副爬行动物确实演化出一些非常“进步”的特征,比同时期的真爬行动物适应力更强。比如石炭纪晚期出现的波罗蜥类,是地球上最早的植食性四足动物之一。
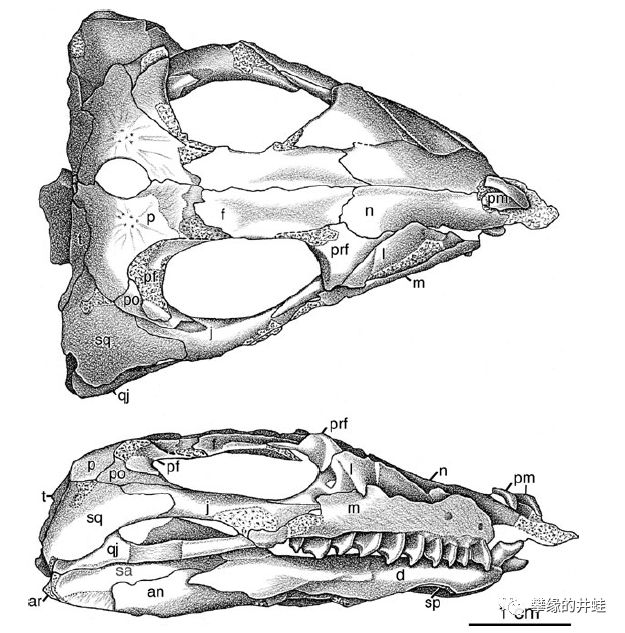
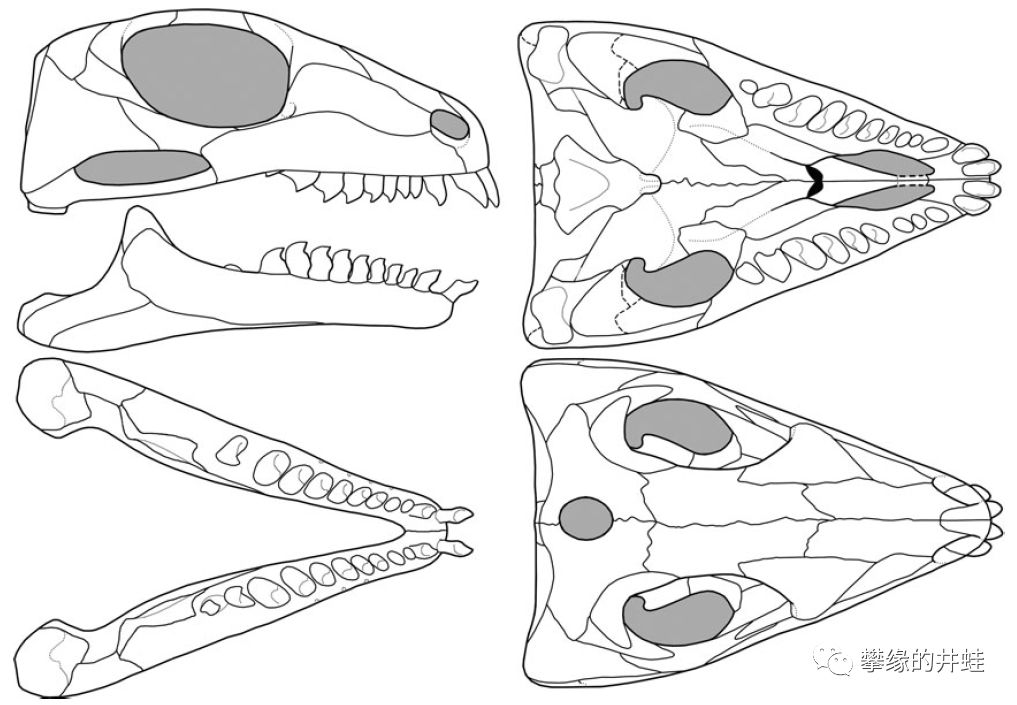
俄罗斯南部发现的二叠纪中期波罗蜥Belebey vegrandis头骨化石和复原图。可以看到它有复杂的牙齿分化,前倾的宽薄“门齿”适合切割植物茎叶,后侧的糙钝颊齿适合磨碎食物,这样的齿型有点类似今天的牛马。另外,这个标本的眼孔下方也有一对颞颥孔,可以让咀嚼肌附着,赋予更强的咬合力。也许就是凭借这些优势,波罗蜥类在二叠纪早期到中期分布非常广泛。图片来源自[1]。

在甘肃玉门也发现了一种二叠纪中期Belebey属波罗蜥:B.vegrandis。化石为右侧上下颌骨,牙齿的特征非常明显。它是俄罗斯的B. vegrandis的近亲,表明这类动物已经扩散了到泛古大陆周边的海岛上。它们一直到二叠纪晚期,才被其他食草动物取代。图片来源自[5],标尺长度1厘米(i图为2毫米)。
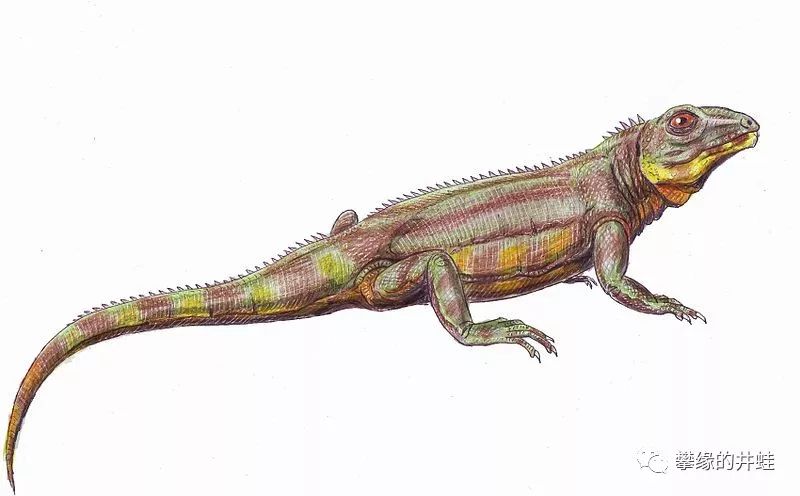
Belebey属波罗蜥复原图。从外表看就是一条蜥蜴,但如果掰开它的嘴,会看到牛马那样的大板牙(没错我们把它抓来研究之后就是这么确定的)。图片来源自Wikimedia。
颞颥孔的数量和位置是判断羊膜动物分类的重要依据(参见第一百九十五回 蛋,爪子,太阳穴和龙兽争霸),但也并不绝对。比如上图中,许多副爬行动物的头骨上都有一对下位的颞颥孔,这应该是和合弓纲的趋同演化。图片来源自[6]。
有一种波罗蜥需要特别介绍一下,那就是生活在2.9亿年前二叠纪早期的真双足蜥Eudibamus cursoris。顾名思义,它可以用两条后腿直立奔跑。这是目前发现的最早采用这种运动方式的四足动物。

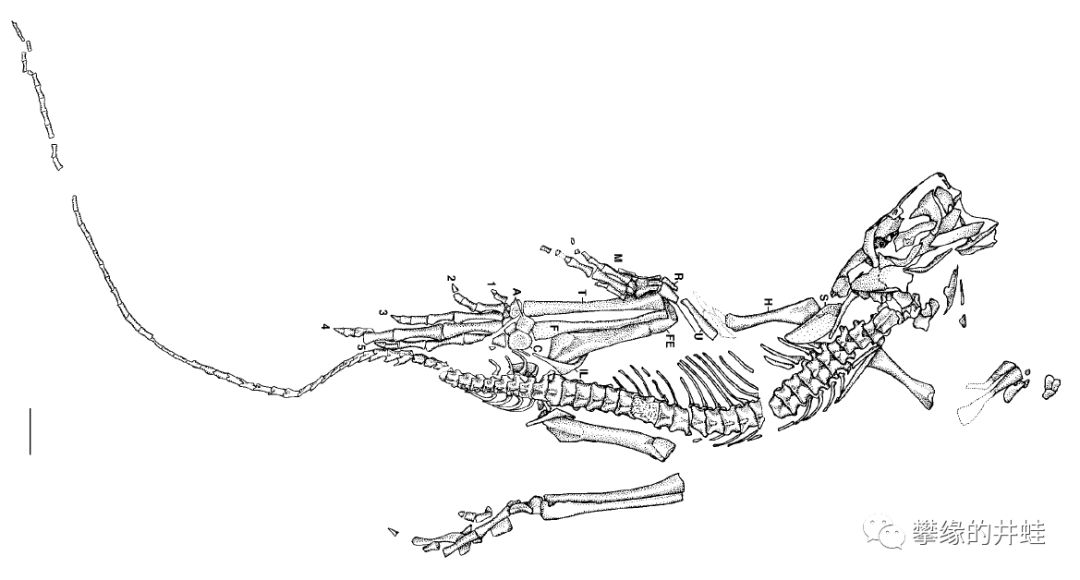
德国发现的真双足蜥化石和复原图(图片来源自网络)和骨骼结构(图片来源自[7])。它的后肢非常发达,比前肢长得多。根据髋骨和足趾的结构,研究者推测它用两条后腿快速奔跑。
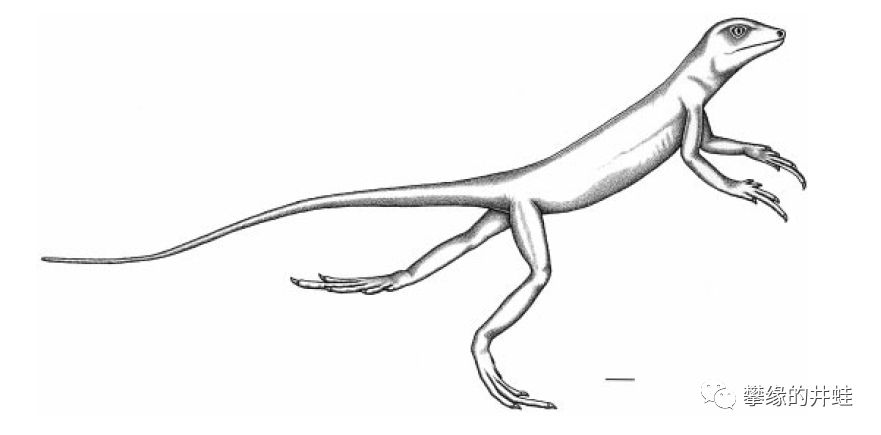
真双足蜥奔跑姿势复原图,图片来源自[7],标尺长度1厘米。作为双足奔跑的先行者,它的结构配置还是显得原始而简陋。后肢不够强健,头颈不够灵活,尾巴单薄,难以平衡体重,还有那一双外八。无常的命运并没有给真双足蜥改善这些缺陷的机会,倒是后来的几类真爬行动物接过了演化的接力棒,把双足奔跑发展到了极致。

真双足蜥的复原图显然参考了一些现代蜥蜴的奔跑姿势,比如上图中的蛇怪蜥蜴(Basiliscus basiliscus)。它们在遇到危险时可以短暂地双足飞跑,甚至表演“水上漂”的绝活儿。二叠纪早期的真双足蜥很可能也用这种方法逃过肉食合弓兽,比如异齿龙的捕杀。图片来源自网络。
再来说说Acleistorhinidae和兰炭鳄科Lanthanosuchidae。这两类副爬行动物也都有一对非常明显的颞颥孔。有的文献把它们放到同一个演化支——兰炭鳄超科Lanthanosuchidea里[6]。
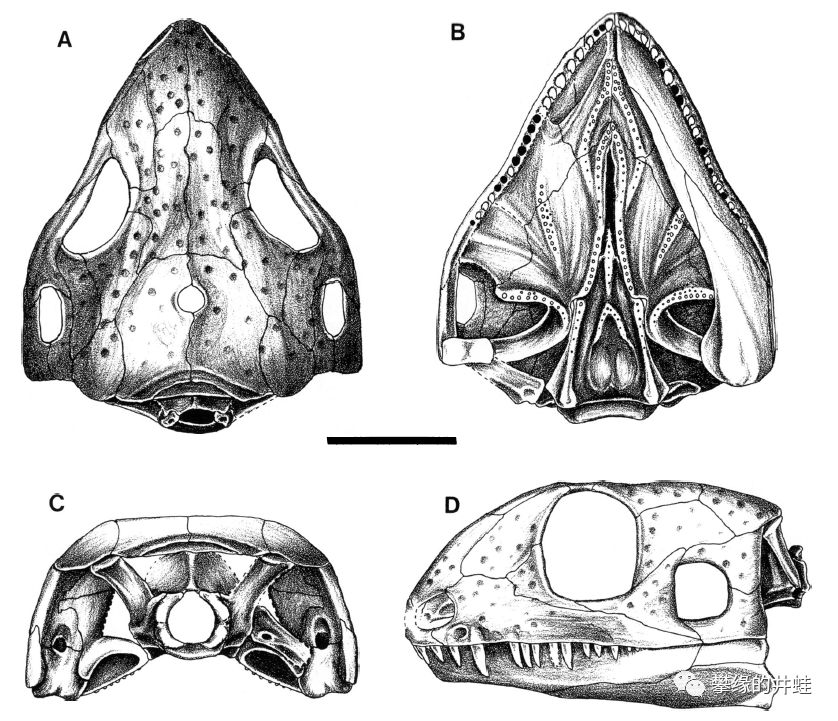
俄克拉荷马州发现的早二叠世副爬行动物Acleistorhinus pteroticus头骨结构,它的眼洞后下方的也有一对颞颥孔。从牙齿上判断,它应该是捕猎昆虫的小型食肉动物。不难看出昆虫的繁盛直接推动了羊膜动物的演化进程。基本上所有的主要演化支都是开局都是食虫的小型种类,然后逐渐演化出中大型的杂食和食草种类,运气好的就会演化出中高级的掠食者,问鼎陆地生态系统的王座。图片来源自[8],标尺长度1厘米。
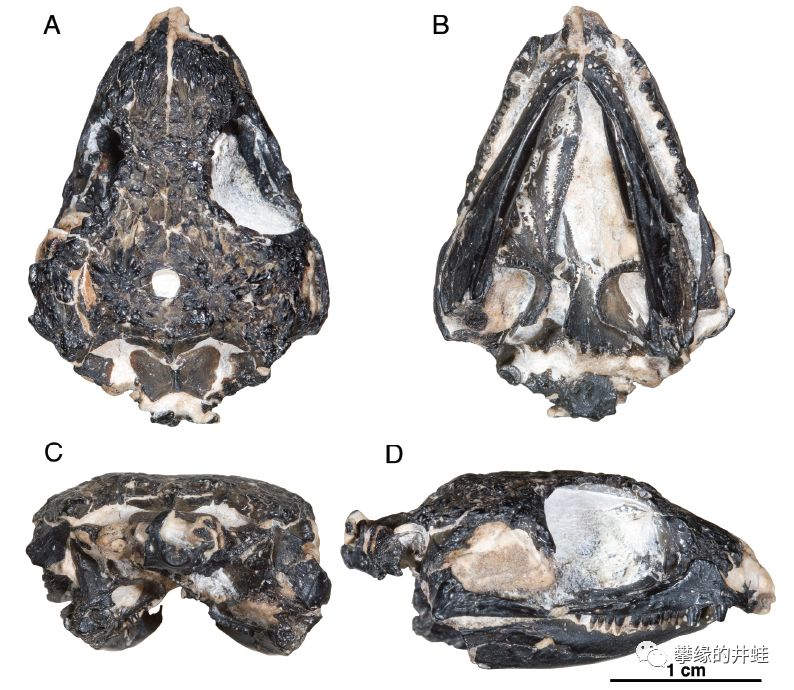
俄克拉荷马州发现的早二叠世兰炭鳄类Feeserpeton oklahomensis头骨化石。它和Acleistorhinus pteroticus确实很相似,尤其是硕大的眼洞和颞颥孔,还有锋利的牙齿。图片来源自[9]。
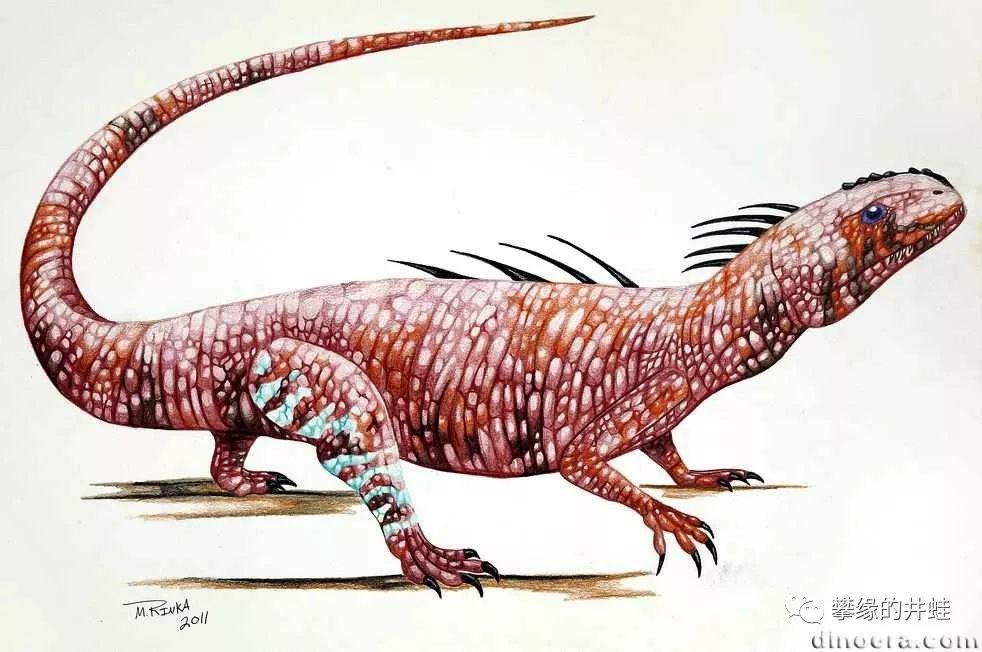
Acleistorhinus pteroticus复原图,图片来源见水印。
Acleistorhinus pteroticus和Feeserpeton oklahomensis是兰炭鳄超科的早期种类,它们明显是轻盈迅捷的小型掠食者。而生活在二叠纪晚期的兰炭鳄属(Lanthanosuchus)则发生了明显的特化,成了一群肥胖笨拙,怪模怪样的家伙。
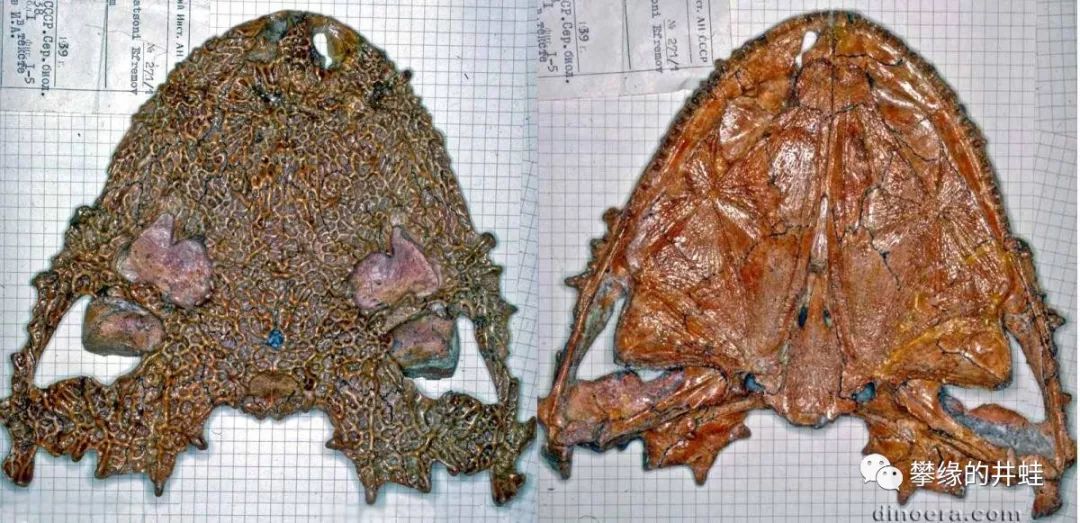

兰炭鳄属代表动物Lanthanosuchus watsoni头骨化石。这种扁额阔口的“蟾头”很像离片椎类。实际上,直到70年代(The Great Soviet Encyclopedia (1979)),兰炭鳄属还被当做古两栖动物。但眼洞后面的颞颥孔却是羊膜动物的铁证。图片来源自网络。
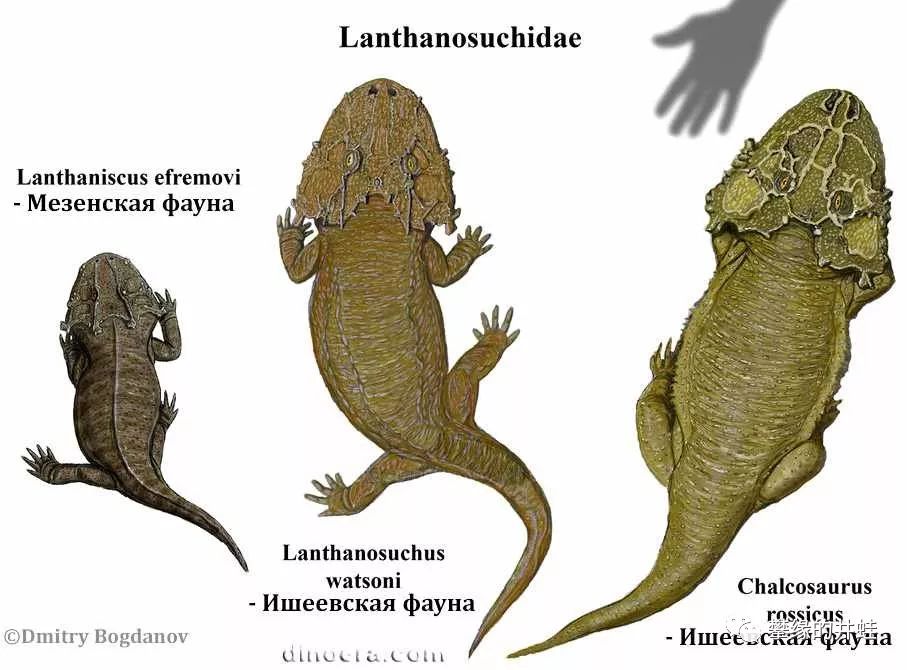
三种兰炭鳄属动物复原图。化石全部发现于俄罗斯鞑靼斯坦,最大的可以长到75厘米长。关于兰炭鳄的资料很少,许多问题都悬而未决。比如它们头骨上别致的纹路是干什么的;比如它们的生活习性是怎样的。这个大扁脑袋无论吃草还是吃肉,都不适合在陆地上进食。许多研究者认为它真的是一种水栖或者半水栖动物,捕食鱼虾,伏击岸边的小动物,也算对得起名字里的“鳄”字。如果真是这样,那兰炭鳄就获得了一项成就:最早和离片椎类抢夺河岸杀手这个生态位的羊膜类。图片来源见水印。
剩下的篇幅,说说前棱蜥类(Procolophonoids)。前棱蜥类也保持着一项成就:唯一熬过二叠纪末大灭绝的副爬行动物类群。
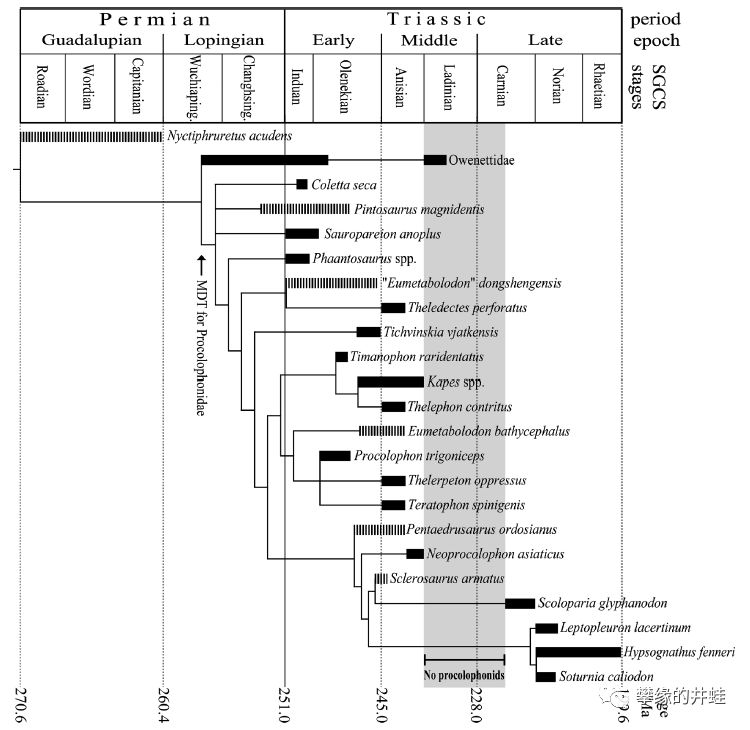
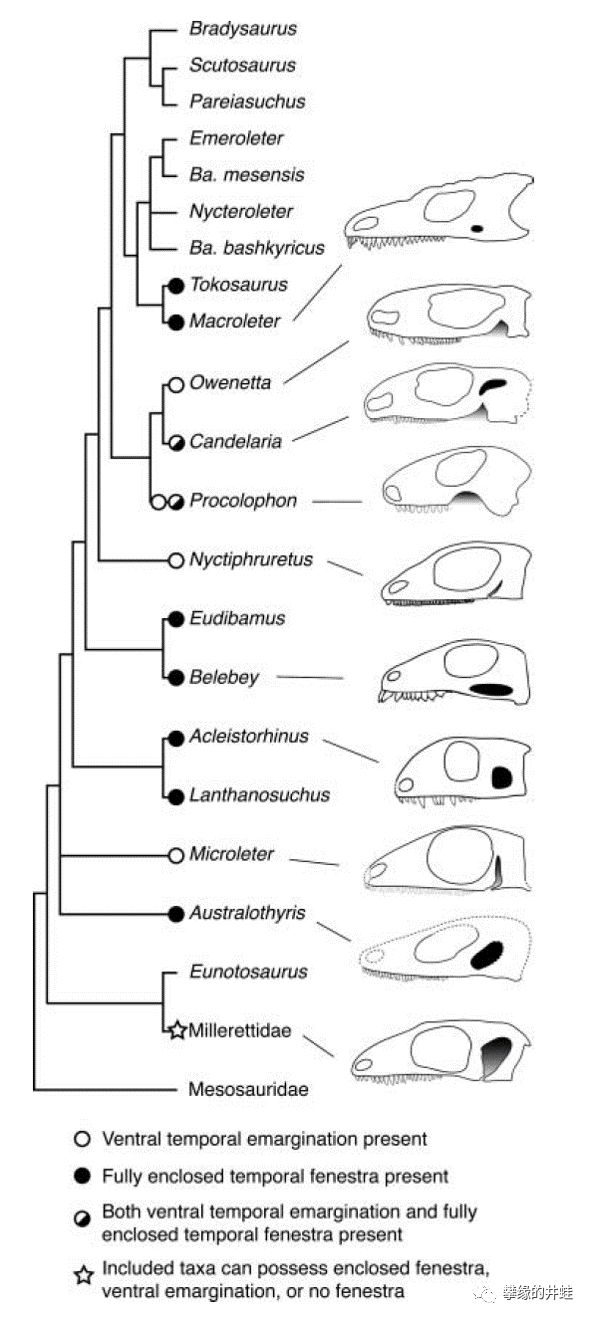
棱蜥类的演化谱系和生存年代。这张图片有几点可以关注一下:最原始的欧文蜥科(Owenettidae)在二叠纪晚期才出现。在三叠纪早期,前棱蜥类一度兴盛,涌现出许多新种类。然后在三叠纪中期出现了一段化石记录的空白(图中阴影部分)。在这段空白之后,大部分前棱蜥类销声匿迹,只有寥寥几种化石出土。最终在三叠纪末的又一场大灭绝中彻底消亡。图片来源自[10]。
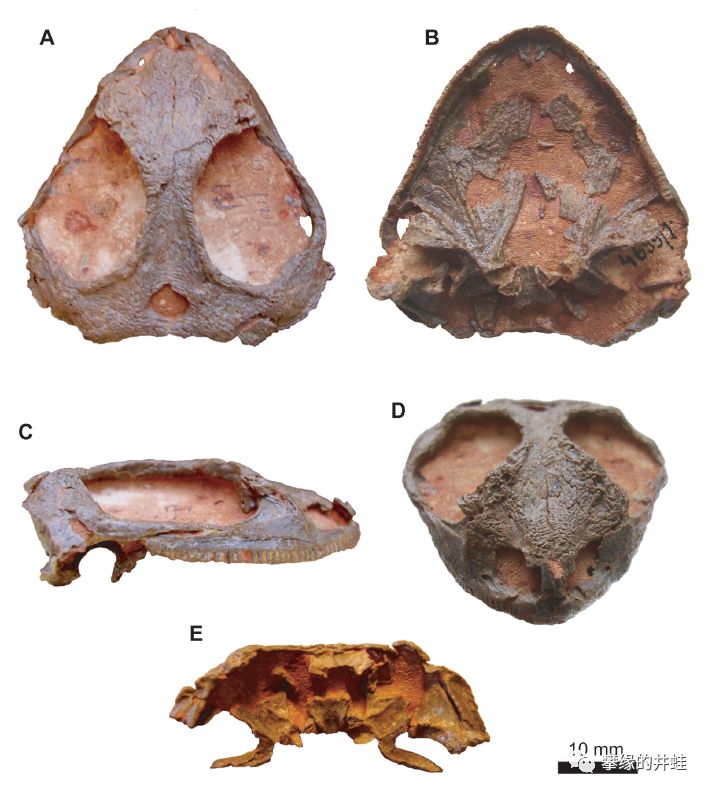
前棱蜥的祖先可能和二叠纪早期的夜守龙属(Nyctiphruretus)亲缘关系较近。上图是俄罗斯发现的夜守龙Nyctiphruretus acudens头骨化石。看上去就是长相普通的小蜥蜴,但是眼洞特别大,表明它的视力不错,也许在夜间捕食昆虫。夜守龙的眼洞后面有一对很小的颞颥孔(注意看C图),这个特征把它和波罗蜥,兰炭鳄联系起来。其他前棱蜥类群则把颞颥孔完全退化掉了,在那个位置长出了别的东西。图片来源自[11]。
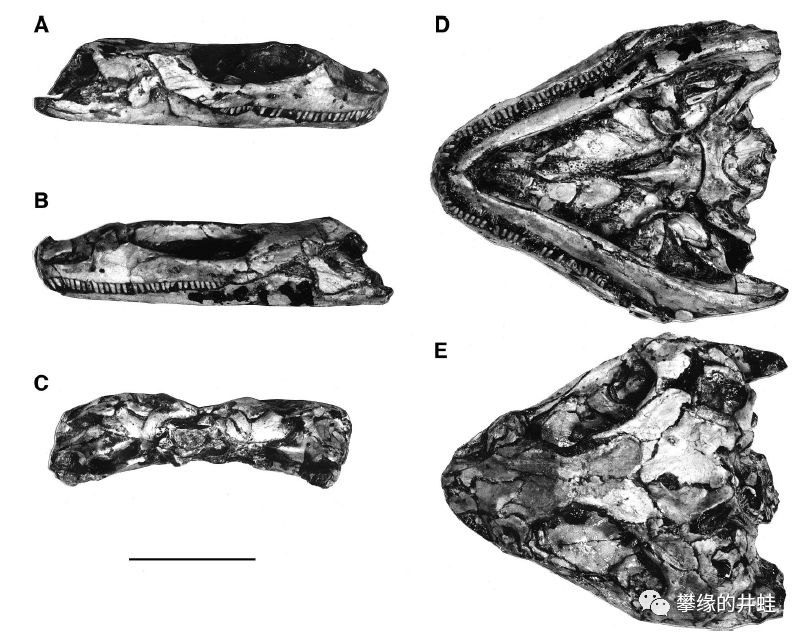
南非发现的晚二叠世欧文蜥Owenetta rubidgei头骨化石。结构很接近夜守龙,也有许多细碎的小尖牙和超级大眼洞,但颞颥孔已经消失了。图片来源自[12],标尺长度1厘米。
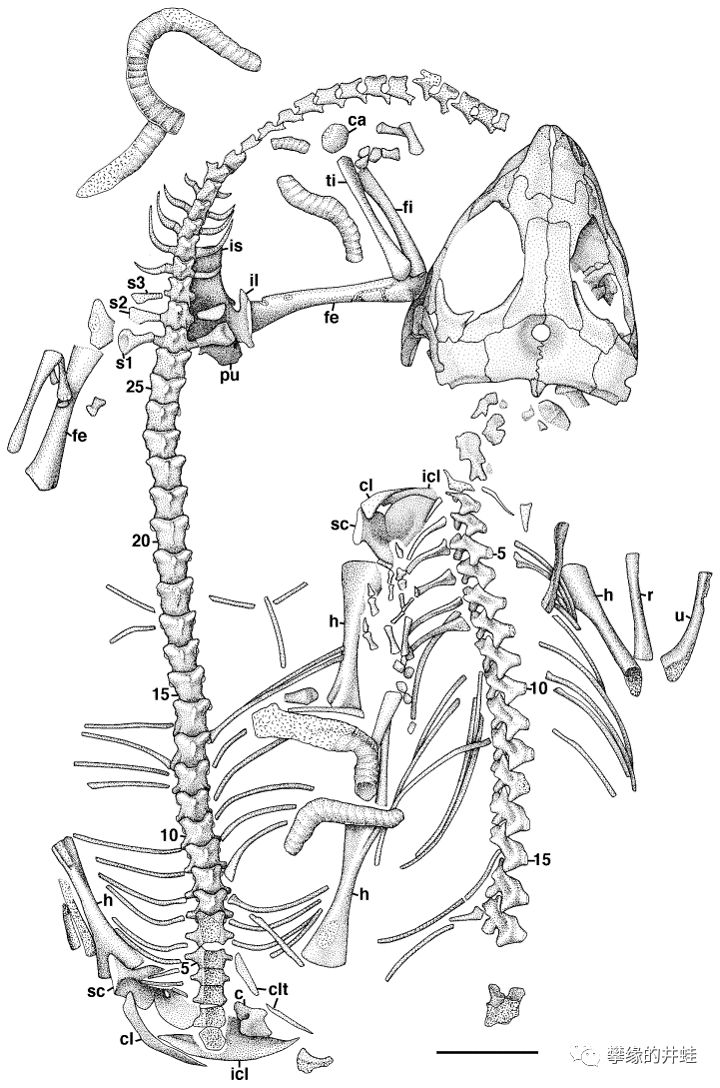

南非发现的另一种三叠纪早期欧文蜥Owenetta kitchingorum骨骼化石简图(图片来源自[12],标尺长度1厘米)和复原图(图片来源自Wikimedia)。看上去就是一条蜥蜴的样子。
夜守龙和欧文蜥处于前棱蜥超科中比较基干的位置,特化不多,长得可以说中规中矩。而似乎是为躲过超级灭绝事件狂欢庆祝一样,三叠纪出现的其他前棱蜥类群开始放飞自我了。
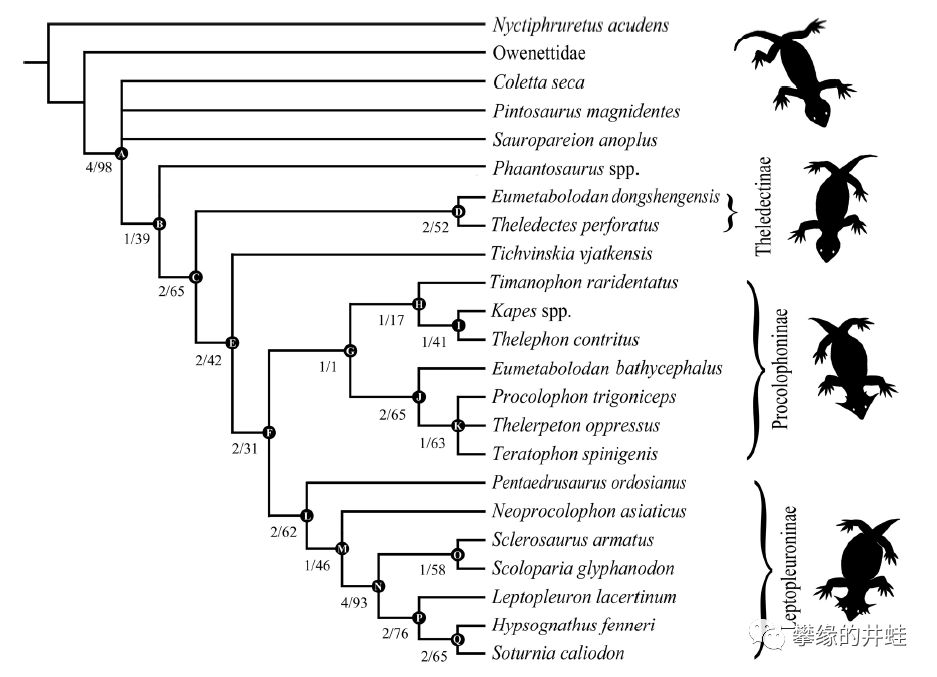
前棱蜥超科主要类群和演化关系。注意右边的剪影的变化。图片来源自[10]。
演化后期的前棱蜥类体型变大,从猎食昆虫转为杂食或者植食,还配上了圆滚滚的草包肚子。最明显的变化是它们的头部出现各种奇怪的装饰,原本颞颥孔所在的颊部骨骼延伸变形,还长出了棘刺之类的附着物。
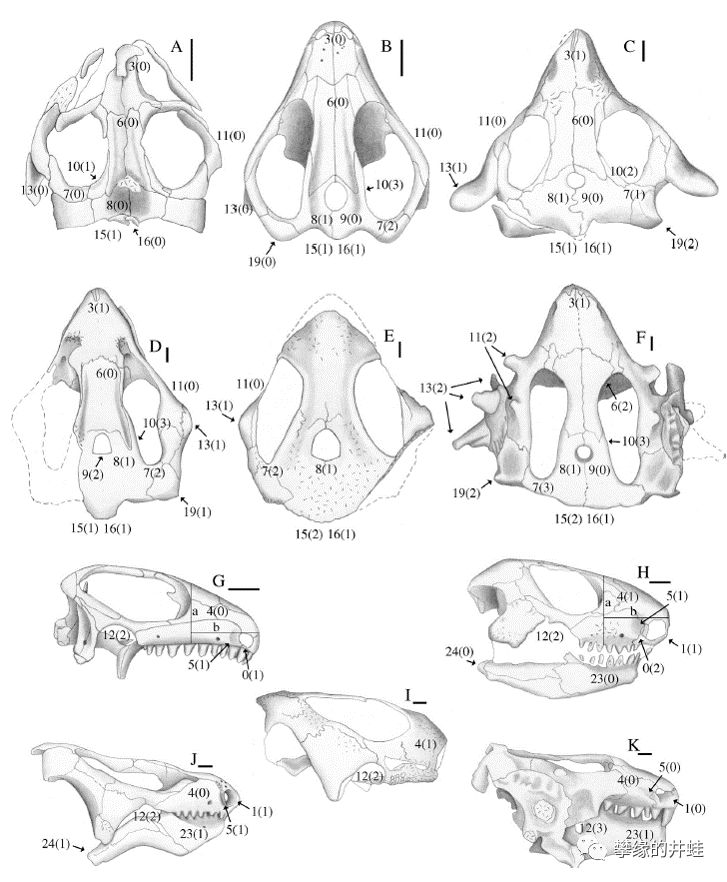
一些世界各地出土的三叠纪前棱蜥类头骨结构。A:Barasaurus besairiei(三叠纪早期,南非卡鲁盆地);B,G:Tichvinskia vjatkensis(三叠纪早期,俄罗斯);C,H:Procolophon trigoniceps(三叠纪早期,巴西南部);D,J:河套五角蜥Pentaedrusaurus ordosianus(三叠纪早期到中期,陕西府谷);E,I:亚洲新前棱蜥Neoprocolophon asiaticus(三叠纪中期,山西榆社);F,K:Hypsognathus fenneri(三叠纪晚期,新泽西州和康涅狄格州,最后的前棱蜥)。在哪个神秘空白出现之前,前棱蜥类仍然非常成功,到处都能发现它们的踪迹。图片来源自[10],标尺长度0.5厘米。

德国巴登(Baden)地区发现的三叠纪中期前棱蜥Sclerosaurus armatus化石。这个标本保存了近乎完整的脊柱和肋骨,一侧肢体,还有部分头骨。它的全长大约半米左右,身体粗壮,腹部膨大,一看就是个四足草包。图片来源自[13]。
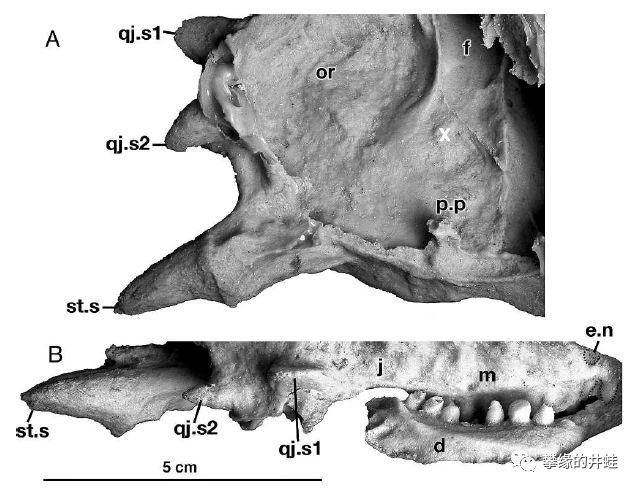
Sclerosaurus armatus头骨部分放大。在它的头部两侧,各有几根角状的长棘(qj.s1,qj.s2,st.s)。图片来源自[13]。

Sclerosaurus armatus复原模型。这么一个半米长的“怪蜥蜴”,还是有点唬人的。图片来源自[14]。
最后的前棱蜥,也是最后的副爬行动物,是生活在三叠纪末期的Hypsognathus fenneri。在它生存的年代,合弓纲已经走向穷途末路,镶嵌踝类主龙经历了短暂的兴起。当副爬行动物的最后一丝余晖消逝之时,恐龙帝国的朝阳正冉冉升起。

Hypsognathus fenneri模式标本化石。它的体型比前辈缩小了一点,继承了扁平浑圆的身体,头部两侧的“角”数量更多,也更加突出。图片来源见水印。

Hypsognathus fenneri头骨化石复原模型。它也有从夜守龙开始一脉相传的大眼洞,但是被拉成了很奇怪的形状。图片来源见水印。

Hypsognathus fenneri复原图。它头部两侧的长棘用途不明,看上去似乎起不了什么防御的作用,也许是某种第二性征,用来同类相争或者吸引异性。图片来源见水印。
除了上面介绍的之外,还有两个非常重要的类群:中龙类和巨齿龙类。它们是副爬行动物演化的巅峰,都在某种程度上突破了“蜥蜴形体”的限制。欲知详情,请看下回分解。
地球名片
生物分类:动物界-脊索动物门-脊椎动物亚门-四足形类-羊膜类-蜥形纲-副爬行动物
存在时间:石炭纪晚期至三叠纪晚期
现存种类:无
化石种类:未找到完整统计
生活环境:陆地,淡水?,内陆盐湖或浅海?
代表特征:分不到真爬行动物的蜥形类
代表种类:波罗蜥类,前棱类,巨齿龙类,中龙类,兰炭鳄类,夜守龙类,等