
前言
治疗性抗体是目前生物制药中最成功的领域。在过去的30多年里,90+种治疗性抗体被批准用于各种适应症,包括癌症、自身免疫性疾病、传染性疾病和心血管疾病。从2014年Blincyto上市获得成功以来,具有与多个表位结合能力的双特异性抗体(bsAb)形式在多方面疾病治疗中显示出广阔的应用前景。然而,由于不同抗体的重链和轻链(HCs和LCs)的混杂配对,bsAb的制备工艺一直具有挑战性。通过随机配对,最多可以有10种不同的组合,其中只有一种是正确组装的bsAb。这个问题限制了bsAb的生产的数量和质量,从而无法及时满足临床前和临床开发的需求,成为bsAb开发过程中致命瓶颈。
研究者们已经开发了许多策略,以解决这一问题。抗体框架改造已被用于改善链错配问题,以促进HC或LC的异二聚化,包括knobs‐into‐holes技术和crossMab技术。尽管取得了一些进展,但是单靠框架设计并不能完全解决问题,抗体生产工艺的广泛优化仍然是必需的。一种解决方案是采用分离表达半抗体,然后在体外组装以生产bsAb。然而,这种方法耗时且成本高昂,需要生成两个不同的稳定细胞系,而且需要各自独立生产。因此,在单个细胞系中产生bsAb仍然是一个很有吸引力的选择,但这种方法需要更广泛的早期优化,以获得能够以最少的产品相关杂质产生最高水平正确组装的bsAb的细胞系。
KOZAK序列与翻译效率
改变翻译效率是一种系统的方法,可以用来控制重组蛋白在哺乳动物细胞中的表达。多肽合成是在翻译起始位点(TIS)开始的,这是一个由起始密码子及其相邻碱基组成的区域。在真核生物中,翻译起始通常遵循扫描机制模型。该模型假设核糖体预起始复合物由40S核糖体亚基Met-tRNA和若干起始因子组成,在信使RNA(mRNA)的5′端结合,并在3′方向线性前进,寻找起始密码子。将小40S核糖体亚基定位在起始密码子后,起始因子分离,大亚基与小40S核糖体亚基结合形成核糖体复合物并开始翻译。
然而,起始并不总是局限于最接近5′端的起始密码子(ATG)。如果第一个ATG密码子出现在一个最佳的环境中,核糖体复合体通常会启动翻译,但是如果第一个ATG三联体周围的TIS是次优的,一些40S亚单位可能会绕过该位点并进一步启动下游的最优表达。因此,细胞可以通过调节TIS来控制蛋白质翻译水平,使起始密码子周围的序列在翻译效率的启动和增强中起主要作用。

Kozak曾报道CCRCCATGG(R=A或G)是一种高效的哺乳动物TIS。在该序列中,位置-3的嘌呤(ATG密码子上游的三个核苷酸)在脊椎动物信使RNA中高度保守。点突变研究提供了证据,证明了位置−3的A或G和位置+4的G(紧随ATG密码子)对于最佳翻译效率至关重要。Kozak序列的长度和核苷酸组成因物种而异,但物种内大多数基因都是保守的。Kozak序列中的点突变会影响高等和低等真核生物的翻译起始,通过改变Kozak序列来调整bsAb亚基生产的翻译强度可以产生更高比例正确组装的bsAb,并减少不需要的错误配对。
Kozak序列突变对翻译的影响
为了证实Kozak序列变体可以通过翻译来调节CHO细胞中的表达,共测试了四个不同的Kozak序列,其核苷酸变异位于Fc融合蛋白起始密码子上游的特定位置。这些变体如图1a所示,预计会降低Kozak序列的平移强度。将含有这些Kozak变体的两种不同的Fc融合蛋白(A和B)瞬时转染CHO细胞,显示CHO细胞中Fc融合蛋白效价降低(图1b)。正如预期的那样,被评估的最强和最弱的Kozak序列变体(分别为CCGCCATGG和CCTTTATGG)的翻译强度与基于Fc融合蛋白滴度的野生(WT)Kozak序列相似或大约一半(图1b)。

图1:Kozak序列突变可调节蛋白表达水平
混合Kozak突变序列的bsAb克隆
为了评价了Kozak序列突变作为一种技术在稳定CHO宿主细胞系中调节蛋白质表达的应用。基于以下标准,选择双特异性抗体Ab1/Ab2作为研究的模型:首先,Ab1/Ab2已具备了一个良好的工艺开发平台,以便于产品的分析和表征。第二,Ab1是一个低表达的分子,在转录下游有瓶颈,而Ab2代表一个高表达分子的例子。因此,这是检验Kozak序列突变在具有挑战性的情况下创建成功的链配对能力的合适场景。第三,Ab1/Ab2作为抗癌药物已显示出强劲的药效,这使得人们对该分子的工艺开发优化更富有兴趣。
此外,Kozak序列选择了4个具有代表性的突变加上WT Kozak进行评估。这些突变的蛋白相对表达强度约为WT的0.3、0.5、0.75、1.3倍。这4个突变涵盖了瞬时转染中观察到的广泛的表达水平。
为了优先促进HC异二聚化,Ab1和Ab2的HCs分别携带了hole和knob突变,电荷导向突变被用于减轻LC错配。将HC和LC结合起来,每一组都有5个Kozak序列变体,每个组共构建25个表达载体。将50个质粒的混合物转染到CHO细胞(称为Kozak mix)。用这种方法产生的多样性可以达到625种不同的链比组合。作为参考值,用WT-Kozak序列克隆的bsAb的四条链也被转染到CHO细胞中(称为WT-Kozak)。采用14天摇瓶来评估每个克隆的产量和双特异性抗体的装配。如图2所示,在两种转染中,单个WT-Kozak克隆比单个Kozak混合克隆表现出更高的抗体滴度。这一结果与实验设计相一致,也符合预期。

图2:kozak Mix和WT Kozak的克隆表达情况
使用非还原CE-SDS评估产品质量,以提供有关分子量形式和其他杂质的信息。该技术可对两种形式进行量化:主峰等同于全抗体(full-ab),包括正确和错误组装的bsAb;以及前峰综合包含低分子量杂质(如半抗体)。在前22个克隆中,WT-Kozak克隆的全ab比例更高(图3)。各克隆的完全抗体百分率各不相同。来自转染2的三个Kozak Mix克隆在60%左右。
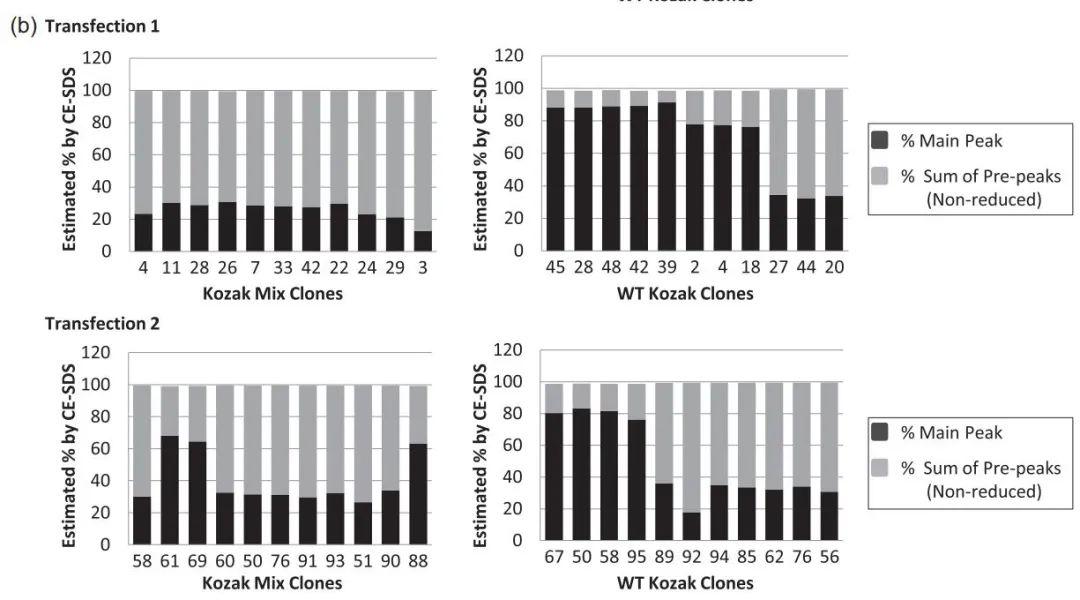
图3:kozak Mix和WT Kozak的克隆表达抗体的质量分析
Kozak序列突变增强了bsAb的组装
根据产量和质量的条件,通过CE-SDS对前4个克隆进行分析,以识别和定量HC和LC的不正确配对。最高的Kozak混合克隆显示40%正确组装的bsAb,这是最高的WT Kozak克隆的两倍多(约18%;图4a)。由于每个Kozak混合克隆可能携带不同的Kozak变体组合,因此每个克隆表现出不同的bsAb组装百分比,而携带相同组合的WT-Kozak克隆表现出更窄的bsAb组装百分比范围。值得注意的是,错误组装的双特异性格式ab2hc+ab1hc+2xab2lc,其LC错配,占WT-Kozak克隆体产生的完整ab的41%-64%。这些数据表明,当使用单细胞系生产时,调节LC翻译水平以获得正确的bsAb组装的重要性。通过质谱对完全抗体形成率最低的Kozak混合克隆(Kozak混合克隆90、11、22、26和28)进行分析,确认这些克隆产生的大多数为半抗体。其形式为Ab1HC+Ab2LC和Ab1HC+Ab1LC。由这些克隆产生的bsAb主要是错误组装的2x ab1hc+2xab2lc,也有2xab1hc+ab1lc+ab2lc的bsAb。

图4:kozak Mix和WT Kozak克隆的质谱定量分析
一旦确定了每个种类的百分比,就可以计算出有效的bsAb滴度。由于一些Kozak混合克隆比WT-Kozak克隆具有更高的bsAb组装效率,Kozak混合克隆克服了先前所示的一般滴度的不足。每种条件下,最高克隆的有效bsAb滴度相似:最高WT Kozak和Kozak混合克隆分别为0.7和0.6g/L。
重要的是,Kozak Mix克隆产品WT Kozak克隆相比产生更少的相关杂质。从大量的副产品中纯化有效的bsAb具有挑战性。由于一些副产物结构与目标bsAb非常相似,因此它们的去除下游往往以牺牲产量或产品质量为代价。区分感兴趣的产品和这些独特的杂质之间可能需要高成本的制造和复杂的技术。Kozak Mix克隆将杂质和产品相关变体的形成降至最低,证明了这种方法潜在的价值。
总结
在这项研究中确定的导致Ab1/Ab2结果改善的Kozak序列是否可以直接应用于提高另一种bsAb的产量仍然是个问题。如果这些抗体与Ab1/Ab2的抗体有相似的情况,可以预测其他bsAb的类似结果。然而,任何给定bsAb的最佳序列可能需要根据经验来确定,因为由此产生的最佳抗体链比率可能取决于该bsAb的特定表达特征。但无论如何,这种技术已经证明了在生产双特异性抗体方面是有用的,并且可以用于解决各种各样的蛋白质表达问题。
参考文献:
Tailoring translational strengthusing Kozak sequence variants improves bispecific antibody assembly and reducesproduct‐related impurities in CHO cells. Biotechnology and Bioengineering. 2020;117:1946–1960.