

在前两回,井蛙简单介绍了发生在GLB 的峨眉山大火成岩省事件,和发生在PTB 的西伯利亚暗色岩事件,以及它们引起的一连串灾变是如何把陆地和海洋生态系统逼到清零的边缘。这一回,我们换一个角度,聊一聊在二叠纪末,主要生物类群的遭遇和命运。
二叠纪的海洋生物化石相对贫乏,保存了动物软体结构的特异埋藏化石群(Lagerstätte )尤其稀少。那些没有骨骼或贝壳的水母蠕虫等等自然绝难发现。不过,我们仍然可以通过一些蛛丝马迹查知它们的存在。





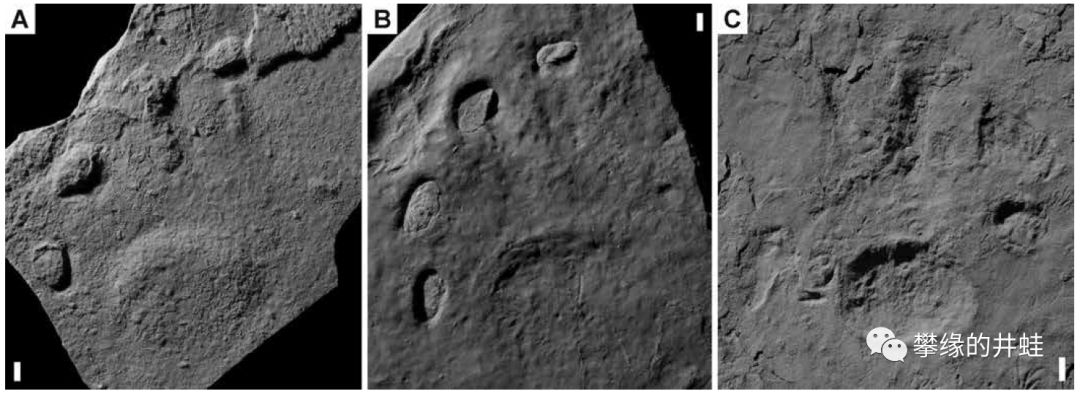
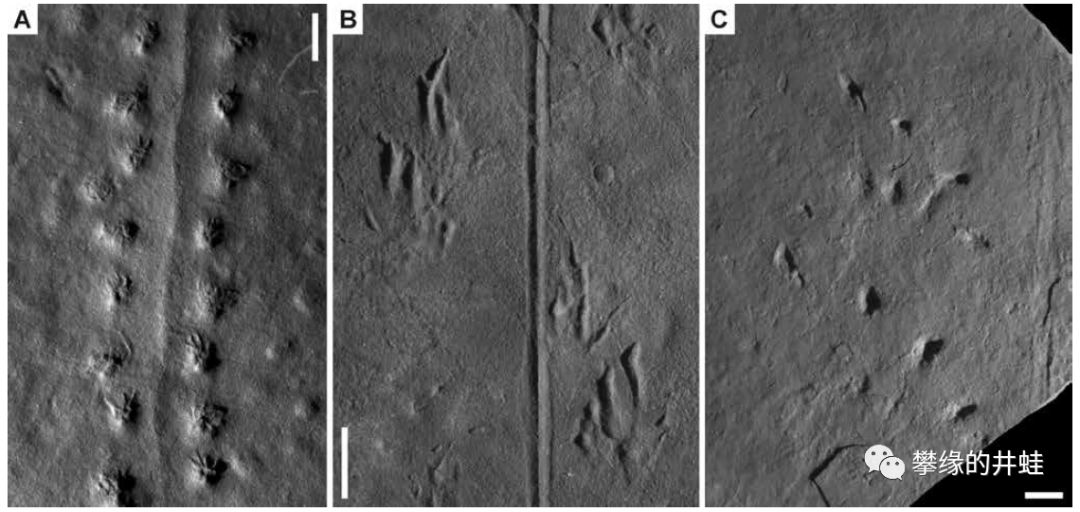
意大利和奥地利交界的普拉莫洛盆地(Pramollo Basin)发现的早二叠世海洋动物足迹化石。在大约3亿年前,这里曾经是位于赤道附近的河流入海口。肥沃的沉积物养活了大量蠕虫,双壳类,腹足类,环节动物,节肢动物,还有其他许许多多的底栖小动物。它们柔弱的身体早已消失得无影无踪,然而留在淤泥上的洞穴和爬痕居然奇迹般地石化保存下来。图片来源自[1]。
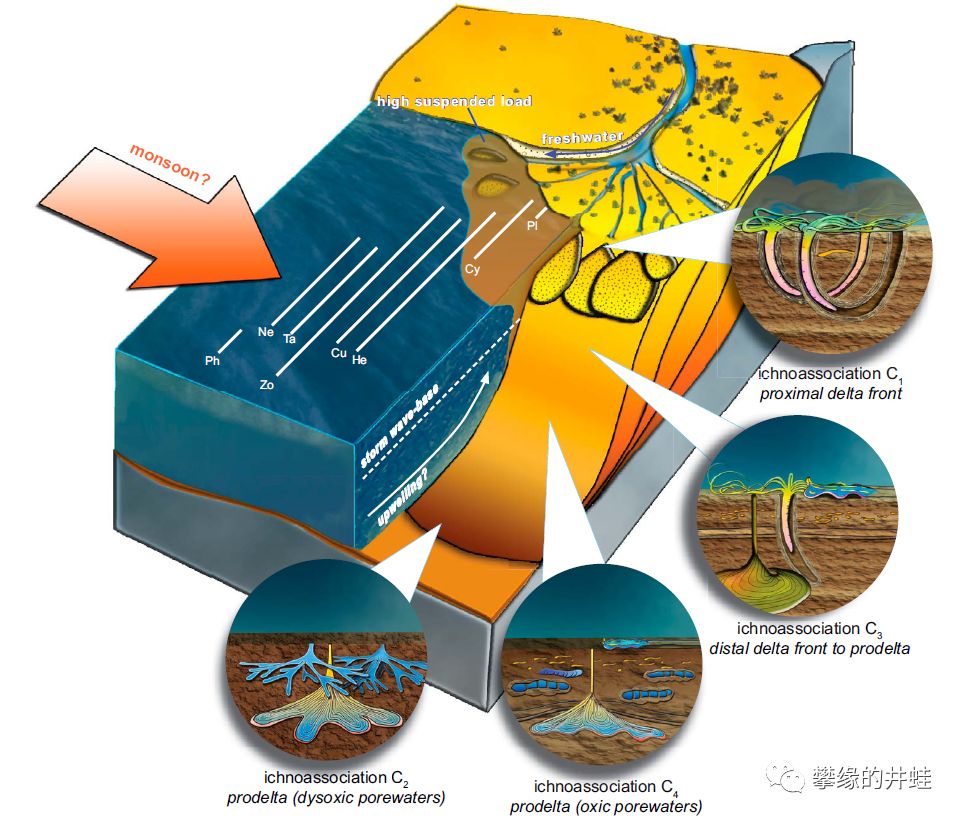
普拉莫洛盆地古地理环境复原图,这种河口三角洲加浅海的环境,很像美国的马荣溪化石群(参见第一百六十一回塔利怪物),连地质年代也很接近(都是大约3亿年前,石炭纪晚期到二叠纪早期)。图中标明了各种足迹化石的分布和生活环境。图片来源自[1]。
有些足迹化石明显是附肢抓挠出来的,可能来自某种节肢动物;有些像是双壳类用斧足顿出来的;有些像是腹足类,石鳖,扁形虫之类用肚子爬出来的。至于那些曲曲弯弯的洞穴,可能性就更多了:扭形动物,环节动物,螠虫动物,星虫动物,鳃曳动物,半索动物,等等,甚至不排除一些不为我们所知的已经灭绝的蠕虫形动物门类。
和其他所有的化石一样,足迹化石也在二叠纪末到三叠纪早期出现断档。这些钻在污泥里的小虫多半都有很强的抗御污染和缺氧的能力,但就连它们也险些被一网打尽。在三叠纪初,各种痕迹化石(包括“洞穴痕迹”和“生物扰动”)一度消失。那些身体柔软的小型底栖动物,就以这种方式反映了大灭绝的规模和影响。
大灭绝摧毁了那些制造礁岩的后生动物。横板珊瑚目(Tabulatecorals )和四射珊瑚目(Rugose corals )灭绝,海蕾纲(Blastoid )灭绝,海百合纲(Crinoid )奄奄一息。海绵和苔藓虫的化石也几乎消失。

澳大利亚悉尼盆地(Sydney Basin)出土的中二叠世棘皮动物化石。A,B为海百合,C为海蕾(部分腕)。也许是曾经的竞争者们让出了生存空间的缘故,幸存下来的少数海百合在三叠纪出现了一轮爆发式的多样化,也算是祸福相倚吧。图片来源自[2],标尺长度1厘米。

另一个在二叠纪末陨落的棘皮动物类群是海座星纲(Edrioasteroid)。这类既像海星又像海胆的小动物在寒武纪就已经出现。二叠纪海座星只在俄罗斯发现了少量化石,全都属于Neoisorophusella属(如上图)。这是这个家族的最后一缕余晖。图片来源自[3]。

悉尼盆地出土的中二叠世苔藓虫化石。图片来源自[2]。苔藓动物门(Bryozoa)是唯一在奥陶纪才出现的主要动物门类(参见第六十六回遗珠)。古生代苔藓虫全部属于窄唇纲(Stenolaemata),其下最繁盛的泡孔目(Cystoporata)和变口目(Trepostomata)在二叠纪末灭绝[4],隐口目(Cryptostomata)残存到三叠纪。现代苔藓虫包括三大类:环口目(Cyclostomata)是窄唇纲的孑遗,大约900种;被唇纲(Phylactolaemata)大约50种,全部淡水生,不产生钙质骨骼,自然也没有化石记录;最繁盛的裸唇纲(Gymnolaemata),大约3000种,最早的化石记录在侏罗纪才出现。像这种演化史被大灭绝一切两半,前后差异巨大的门/纲/目还有很多很多。
以大灭绝为分界,腕足动物和双壳类的地位彻底颠覆,这在之前的回目(第二百十九回寒武纪的余烬:腕足动物篇;第二百二十回嘎啦簸箕翻身记)里已经介绍过。加上软舌螺灭绝[5] ,基本上,触手冠动物在二叠纪之后一蹶不振,沦为底栖滤食动物中的二三流角色。
当然,双壳类也不是轻松渡过大灭绝。上回提到了小型化效应(Lilliput effect ),二叠纪双壳类的体型普遍大于腕足动物,自然也承受了更大的生存压力。如果不是运动能力,滤水系统的优势,双壳类还真不一定能熬得过腕足动物。
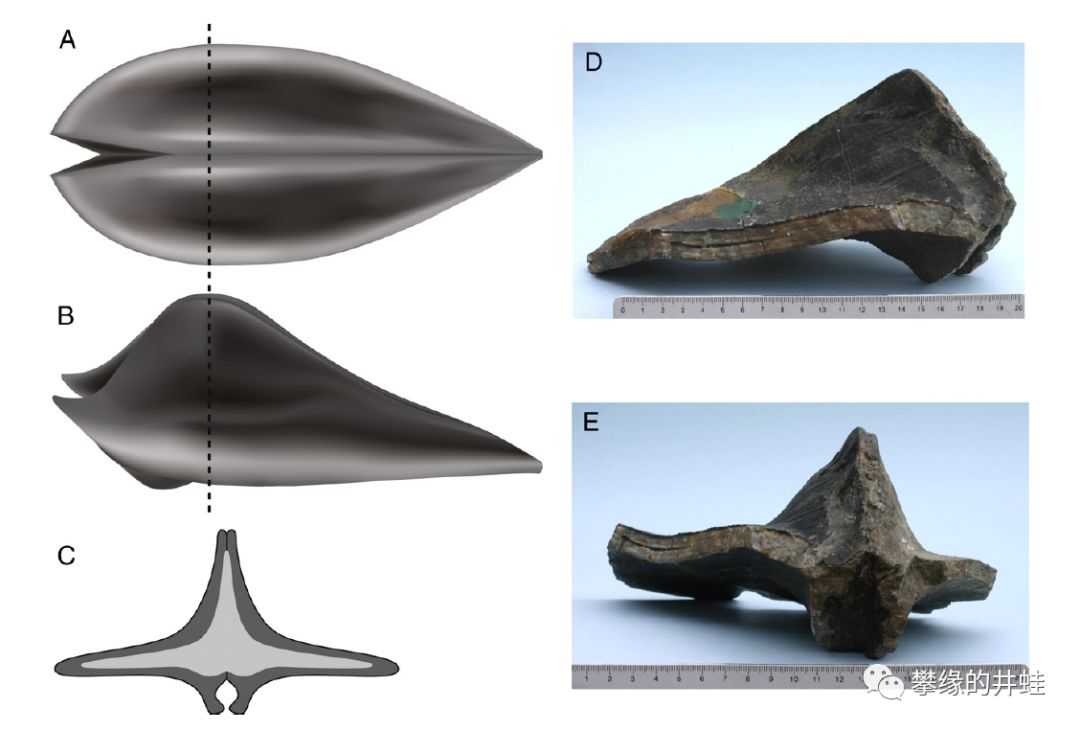
一些二叠纪翅蛤科(Alatoconchidae)的贝壳可以长到1米长,是古生代最大的双壳类。上图是日本发现的巨型翅蛤Shikamaia ogulineci贝壳碎片化石(D,E)和翅蛤科完整贝壳复原图(A-C)。它们的贝壳在水平方向延伸成非常夸张的形状,应该是像今天的砗磲一样,固定在海底不动。图片来源自[6]。
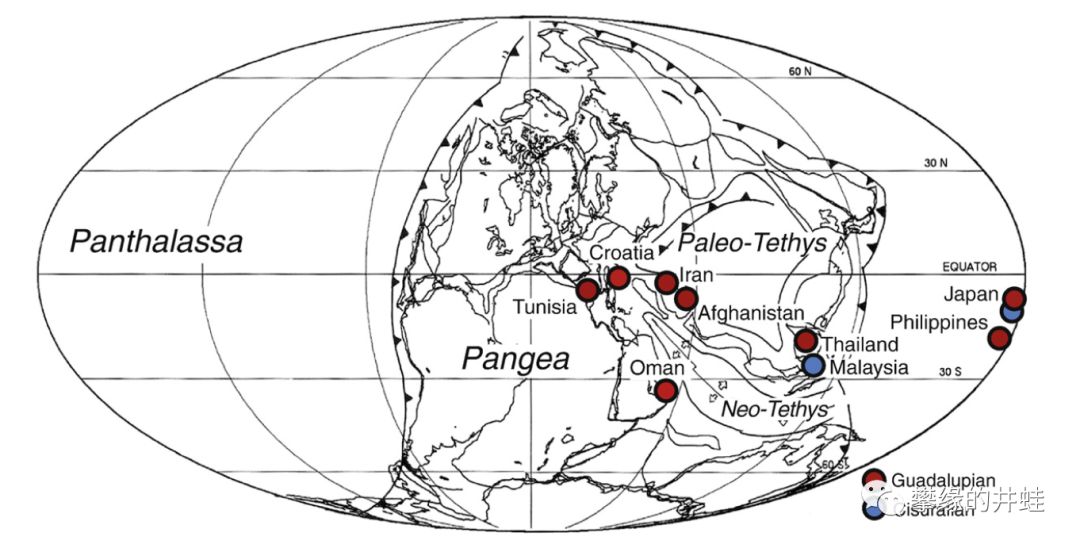
翅蛤科化石分布很广(如上图所示),可见二叠纪海洋的富饶。这些巨大的双壳类在GLB灭绝,它们很可能是峨眉山大火成岩省的受害者。图片来源自[6]。
二叠纪末也是腹足类演化史的一道分界线,在古生代占据优势的类群——古腹足类(Archaeogastropoda 或Vetigastropoda )迅速衰落,包括新腹足类(Caenogastropoda )在内的其他演化支崛起成为螺类的主流。







俄罗斯发现的中二叠世古腹足类化石,看上去和今天常见的螺蛳好像也没什么区别,图片来源自[7]。很可能是因为繁殖方式的原始(没有交配器官,体外受精),古腹足类在大灭绝之后逐渐没落。现存的古腹足类(包括鲍科Haliotidae,裂螺科Fissurellidae等)全部海生,大约3700种[8],占腹足纲(超过10万种)的比重几乎微不足道。
虽然没有翅蛤那么巨大,二叠纪也不乏超过10 厘米的大型腹足类(好像在任何时代,最大的螺都远远比不上最大的蛤)。毫不意外地,这些大块头都被一网打尽,幸存的腹足类都是一些毫米级的小东西。


上图:晚二叠世大型腹足类Pleurotomaria。发现地:日本岩井崎石灰岩(IwaizakiLimestone),图片来源自[9],标尺长度1厘米。下图:中二叠世腹足类Keeneiaocula。发现地:澳大利亚悉尼盆地。图片来源自[2],标尺刻度为厘米。
头足纲中,杆石目(Bactritida )灭绝,直角石目(Orthocerida )挺到了侏罗纪,然后灭绝。残存的鹦鹉螺目(Nautilida )躲到深海养老去了,居然一直苟到了今天。蛸形类(Coleoidea )化石太少不做分析。菊石类(Ammonoidea )只剩下星星点点的火苗。软体动物的适应能力真的很强,除了喙壳纲(Rostroconchia )这个倒霉蛋出局(参见第二百二十一回就是一盘大螺贝),双壳纲,腹足纲,尤其是头足纲在大灭绝过后,无论种类数量还是体型都很快恢复,撑起了海洋食物链。三叠纪早期能突然涌现出那么多的海爬,软体动物居功至伟。
节肢动物的老古董三叶虫和板足鲎灭绝。肢口纲(Merostomata )只剩下剑尾目(Xiphosura )的水瓢们,它们和干瘪的海蛛纲(Pycnogonida )一起苟延残喘了2.5 亿年。甲壳动物接收了空缺的生态位,一直繁盛到今天。
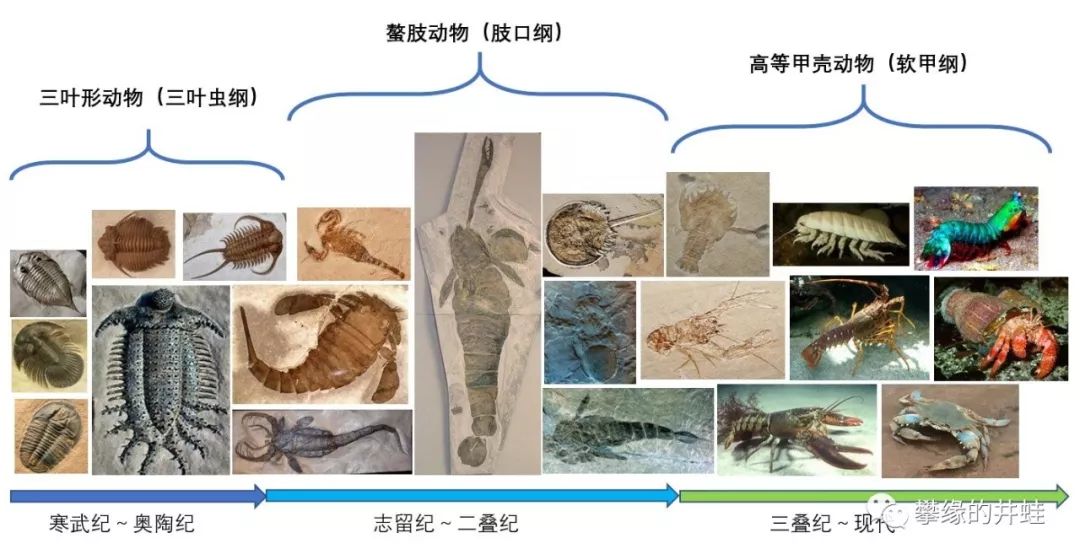
简单做了一张图。海洋节肢动物清晰地显示了三大演化动物群的更替。从寒武纪演化动物群(CambrianEvolutionary Fauna),代表:三叶虫;到古生代演化动物群(PaleozoicEvolutionary Fauna),代表:板足鲎;再到现代演化动物群(ModernEvolutionary Fauna),代表:软甲纲甲壳动物。
软甲纲中,石炭纪到二叠纪极盛的臀头虾类(Pygocephalomorphs ,参见第二百二十二回虾啊,真虾,真是虾,是真虾… )灭绝,我们熟悉的十足目(Decapoda )开始攻城掠地。
古生代十足目甲壳类化石很少。上图是阿肯色州发现的早石炭世十足目甲壳动物Imocaris tuberculata。这些真正的虾兵蟹将演化出大钳子来击破软体动物的贝壳,在三叠纪迎来一波大发展。图片来源自[10],标尺长度5毫米。
有一类微型甲壳动物——介形纲(Ostracoda )留下了非常丰富的化石资料,常被用做划分地层,研究古气候和古环境的重要依据。它们自然也在地层中深深记下了二叠纪末大灭绝的威力。
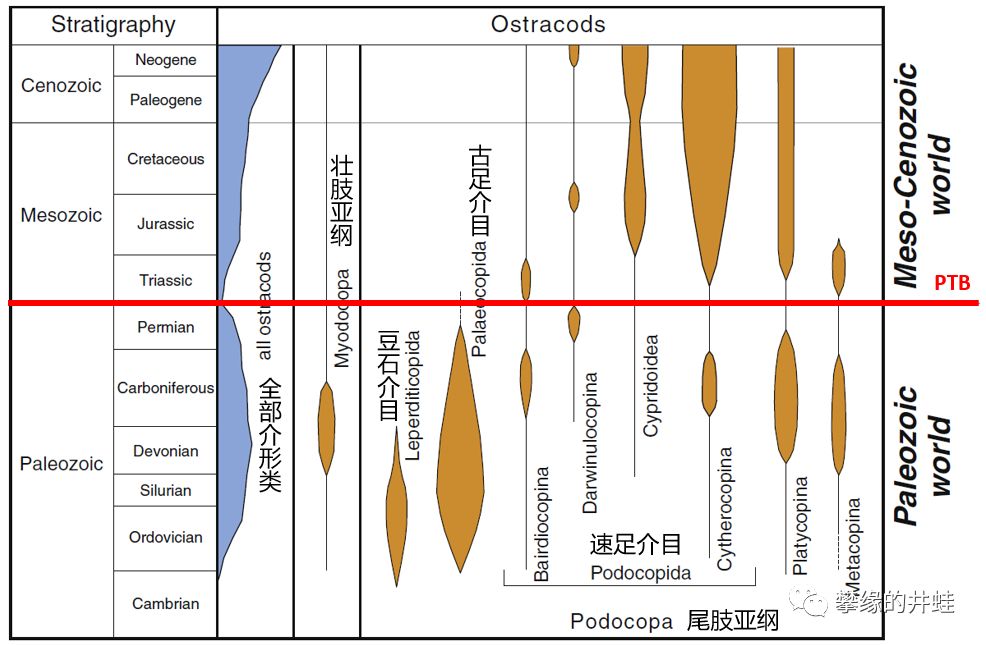
介形类自奥陶纪出现(参见第八十一回芥豆),种类基本保持平稳。唯独在二叠纪末出现一次锐减。古足介目灭绝,速足介目兴起。这个微小的甲壳动物家族,也悄然完成了一场王朝更替。图片来源自[11]。
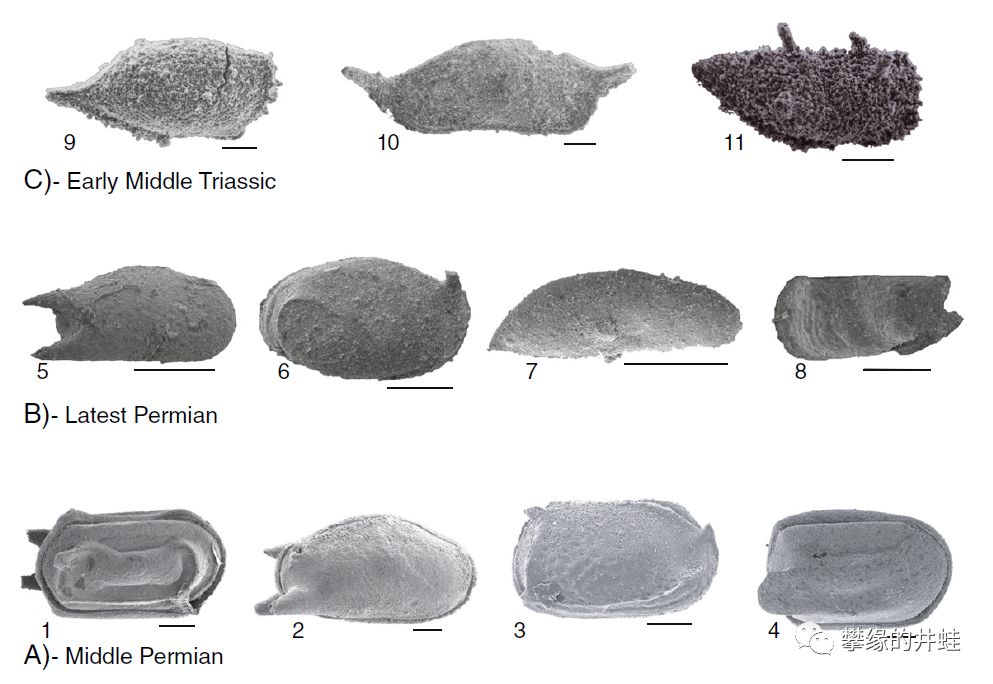
一些二叠纪中晚期到三叠纪早期介形类化石。介形类有左右两片壳瓣包夹着分节的身体和附肢。图上的都是壳瓣化石。图片来源自[11],原文献中并没有给出标尺长度,推测应该是0.5毫米或者1毫米。
类似介形类的微化石还有䗴类(Fusulinid ,参见第一百七十四回 Single Cell )和牙形刺(Conodont )。一个在二叠纪末灭绝,一个在三叠纪末灭绝。

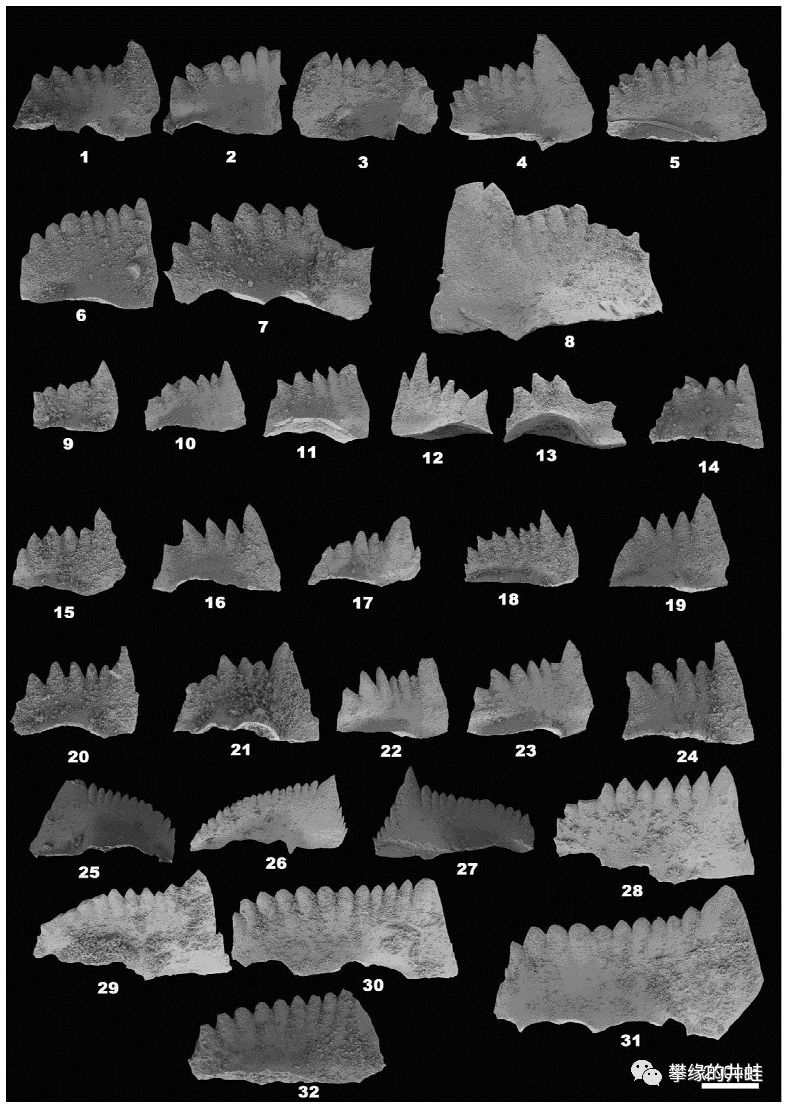
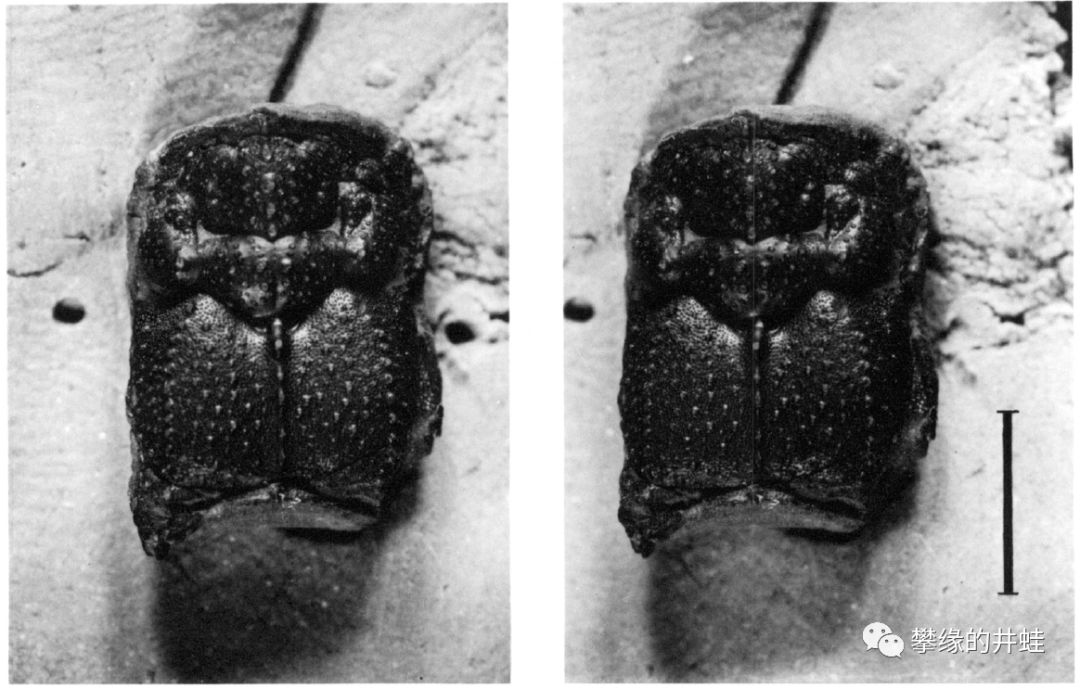
重庆中梁山凉风垭PTB附近的牙形刺。牙形动物(参见第八十九回碎骨狂魔)就是用这些牙齿碾碎像介形类和䗴类的小生物。图片来源自[12]。
二叠纪鱼类化石稀少而且散碎。我们姑且假定那些在三叠纪之后再也没有发现化石的类群,比如古老的棘鱼类(参见第二百二十七回锦鳞记6 :金鱼池畔草蒙茸),各种奇形怪状的全头类(参见第一百八十一回鲛腾记2 :形),长着奇怪头饰的西莫利鲨目(参见第一百八十二回鲛腾记3 :齿)和尤金齿鲨目(第二百二十五回鲛腾记4 :轮),都是大灭绝的牺牲者,大约未必十分错。
大灭绝对硬骨鱼类影响,最明显的就是庞杂的“古鳕类”种类骤减。菊石在三叠纪抢先复苏,占据了相当比例的游泳动物生态位,软骨鱼仍然扮演着中型掠食者,蜥形纲也开始下海,这些压力都迫使硬骨鱼类演化出新的类型。
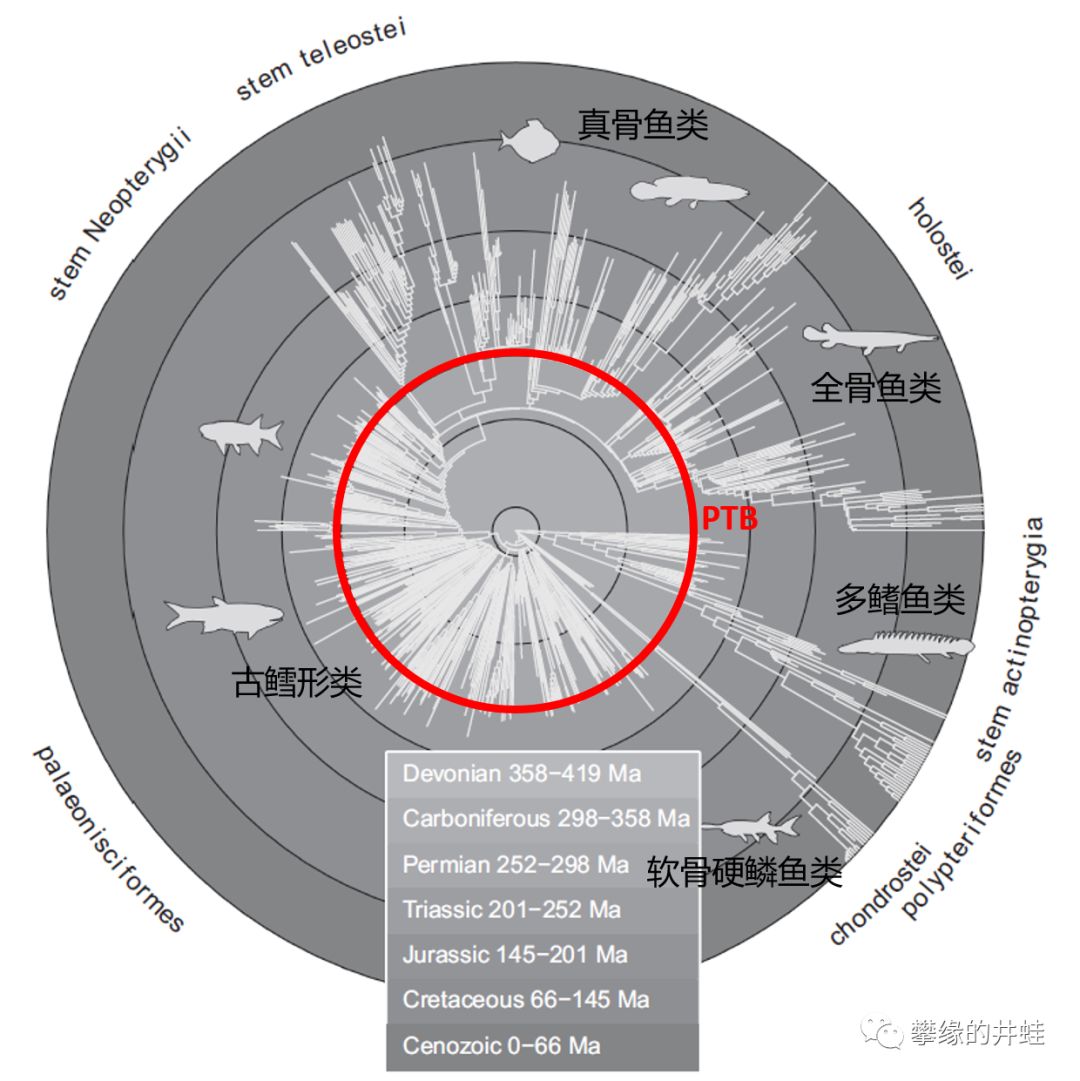
硬骨鱼类主要类群的演化谱系和生存年代。古鳕类在PTB之后迅速衰落,三叠纪末又遭受一次重击,把生态位让给了更快,更灵敏,摄食能力更强的新类群。图片来源自[13]。
再来看看陆地和淡水的情况。
“龙兽争霸”的双方都受到了重创,并将在三叠纪发生地位逆转。
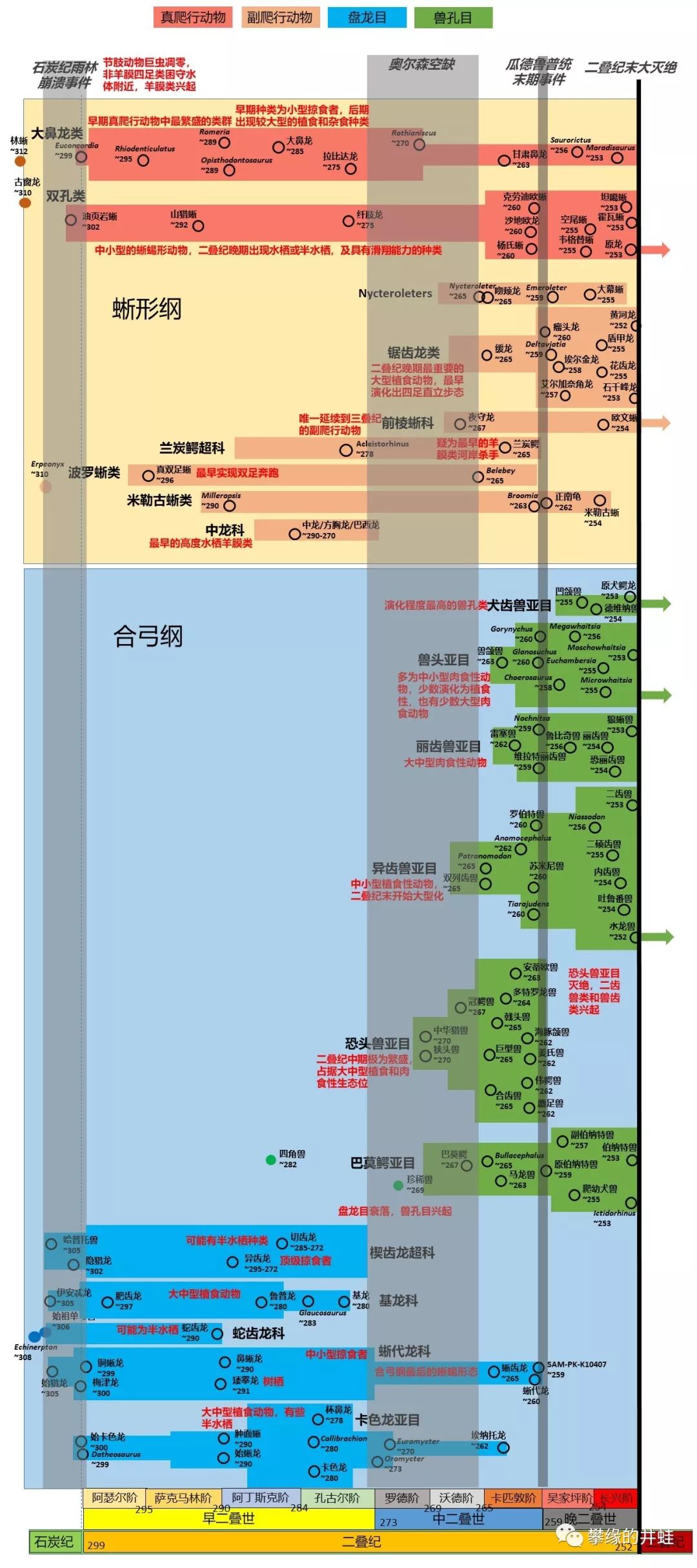
井蛙做了这张图来说明蜥形纲与合弓纲第一回合(古生代)的战况。基龙,杯鼻龙,麝足兽,锯齿龙这样的大草包;异齿龙,巨型兽,安蒂欧兽,狼蜥兽这样的超级杀手,在二叠纪来了,又走了。残存的兽头兽和二齿兽在三叠纪短暂地回光返照之后灭绝,只剩下犬齿兽一支蜷缩在蜥形纲的阴影之下。
壳椎类灭绝(参见第二百三十回螈怪)。残存的离片椎类依然占据着淡水食物链的高层,但它们的时代即将结束,因为蜥形纲即将把河湖水系也纳入治下。滑体两栖类在二叠纪末出现并奇迹般地幸存下来(参见第二百二十九回蟾祖),成为龙兽两族之外的四足动物仅存的血脉。




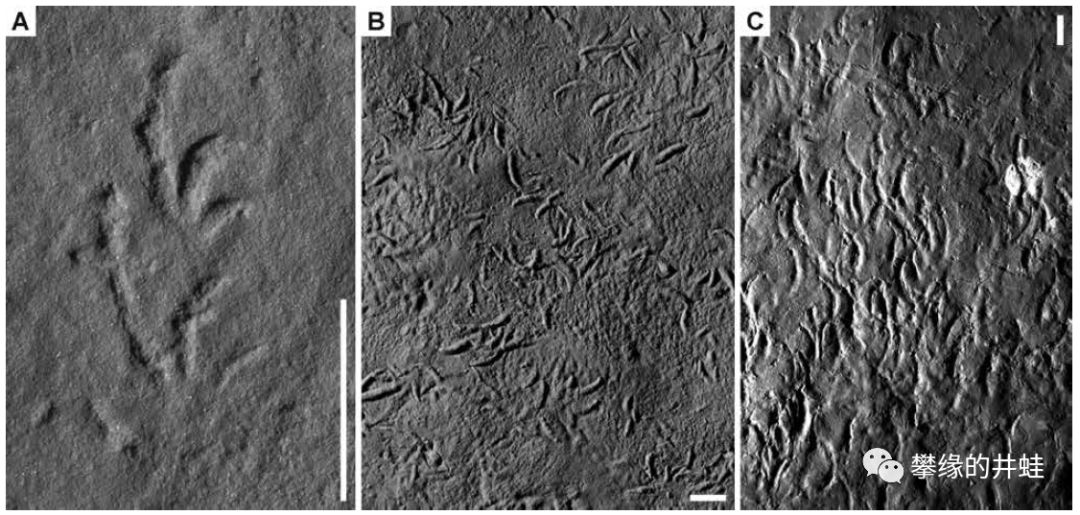
一些早二叠世四足动物足迹化石。留下这些足迹的动物可能包括引螈,双疏螈,鳞鲵,阔齿龙,楔齿龙,蜥代龙,大鼻龙,副爬行动物等等。它们全都没能跨过PTB这条死线。图片来源自[14],标尺长度1厘米。
昆虫在二叠纪末发生了演化史上唯一一次大灭绝。古翅类除蜻蜓目和蜉蝣目外全部灭绝。那些巨大的,凶猛的,张牙舞爪的,好像长着6 只翅膀的怪虫就此绝迹。三叠纪的天空迎来了新的主人,昆虫向着小巧精细的方向的演化。

我国发现的晚二叠世魁翅目昆虫Brodioptera sinensis(应该是叫中华什么虫,井蛙没查到中文名)化石和复原图。魁翅目(Megasecoptera)是一类长着刺吸式口器的古翅类昆虫,在二叠纪末灭绝。图片来源自[15],标尺长度1厘米。
蛛形类也在二叠纪损失惨重,但是和超级地幔柱没什么关系(参见第二百十二回铁虿狰狞12 :二叠纪初大灭绝事件)。
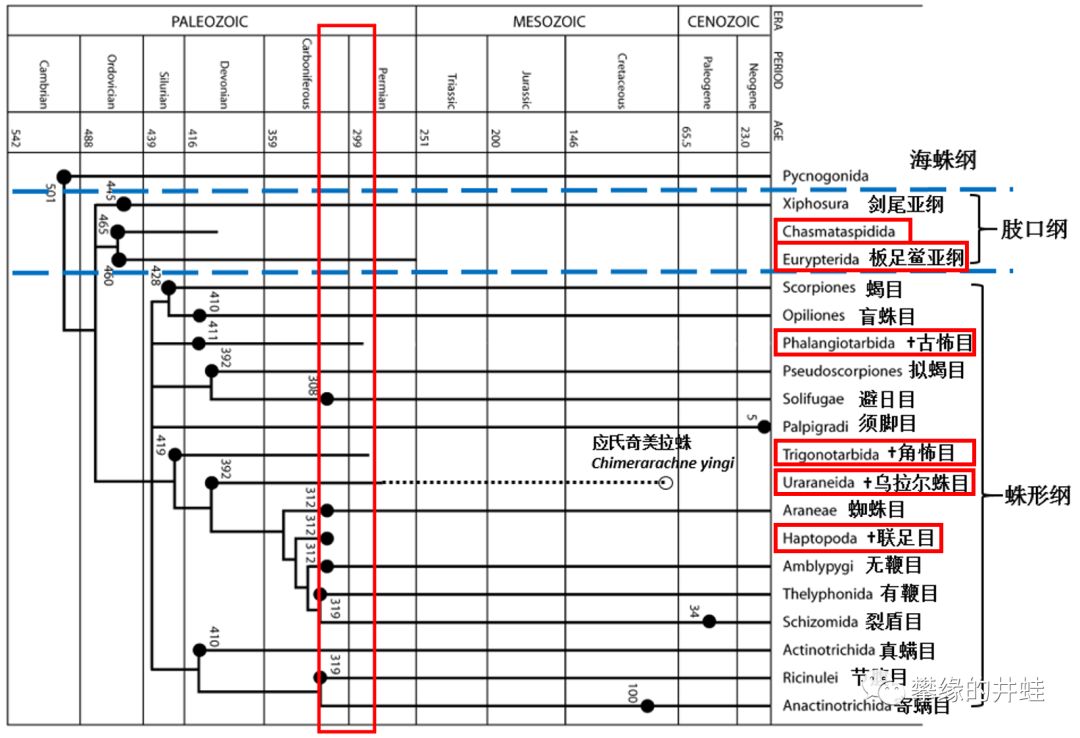
螯肢类主要类群的生存年代。角怖目和古怖目占古生代蛛形纲种类的80%,它们在二叠纪初在消失。在蜘蛛崛起之前,蛛形类乃至螯肢类还要在低谷中徘徊很久。图片来源自[16]。
根据主流统计,陆地生态系统的情况(大约75% 的物种灭绝)比海洋生态系统(大约96% 的物种灭绝)要好一些。这很大程度上是因为陆生植物的坚挺。当然,从雨林崩溃,到超级地幔柱爆发,石松类,节蕨类,真蕨类,种子蕨类和裸子植物的比重也在不断改变。但全部陆生植物的多样性并没有大的波动,它们是灰暗的大灭绝中唯一的一抹亮色。
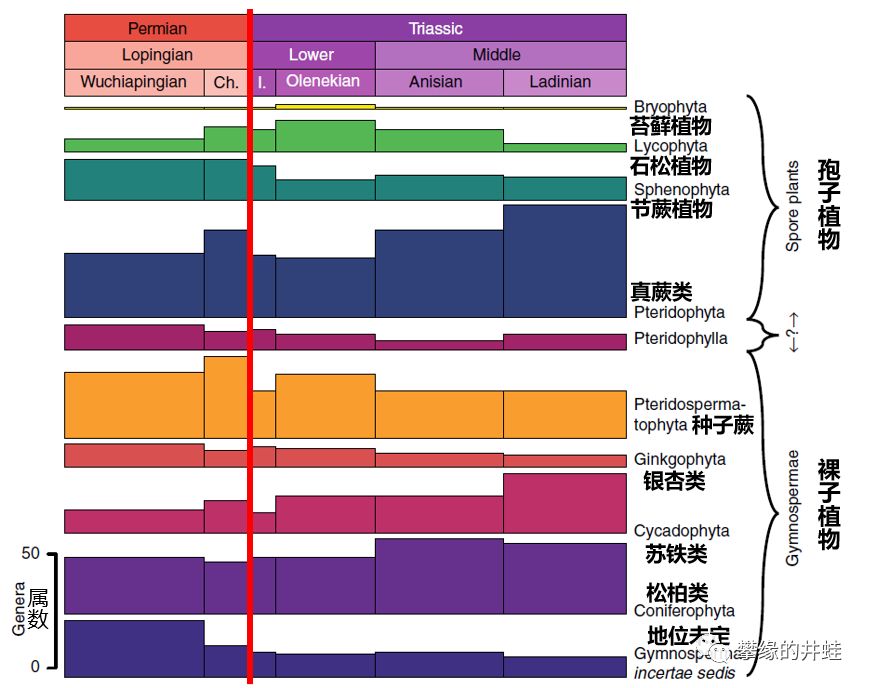
二叠纪晚期到三叠纪陆生植物多样性统计,并没有出现低谷和断层。石松类的衰减,被真蕨类和裸子植物的发展填补,好像只是普通的演替。在二叠纪末的一片哀鸿中,显得非常特别。图片来源自[17]。
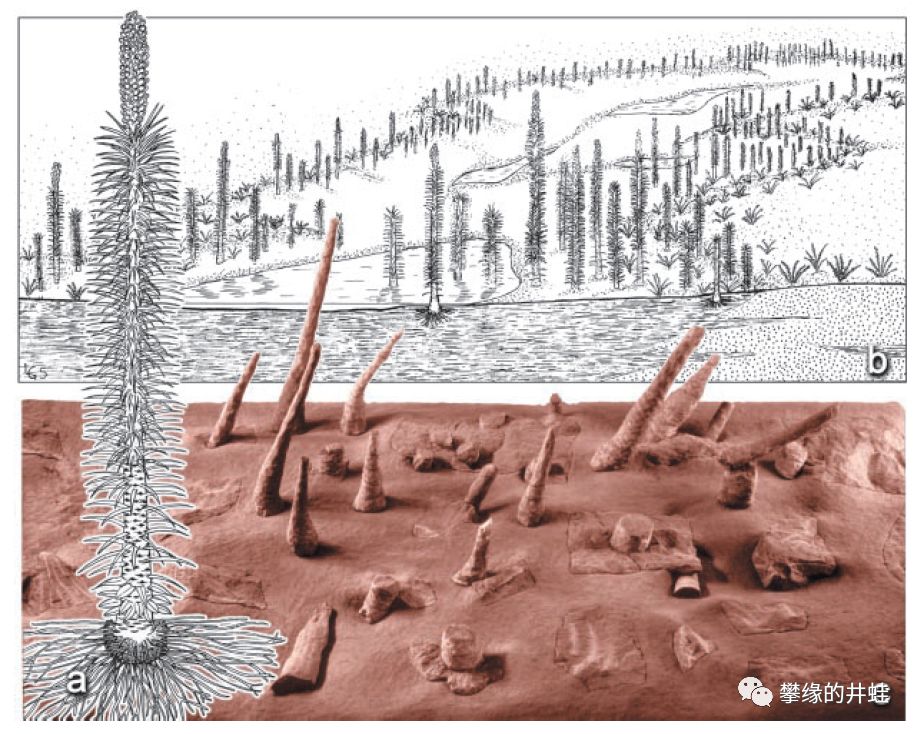
在炎热干旱的三叠纪,低矮的草本植物取代了森林。上图是一类三叠纪早期常见的大型草本石松:1.5到2米高的肋木属Pleuromeia。图片来源自[18]。
有了这片生命的绿色,陆地生态系统就有了复苏和重建的基础。这些顽强的植物支撑着幸存者们熬过长达百万年的灾变,直到生命再次繁荣。

图片来源自网络。
地球名片:
地质时期:显生宙—古生代
起止时间:5.4亿年前至2.5亿年前
持续时间:2亿9千万年
发生事件:生物礁高度发育,菊石繁盛;合弓纲羊膜动物繁盛;昆虫多样化;种子蕨和裸子植物植物取代石松和节蕨;华力西运动完成;西伯利亚暗色岩事件;二叠纪末大灭绝
代表生物:异齿龙,基龙,旋齿鲨,锯齿螈,杯鼻龙,麝足兽,冠鳄兽,狼蜥兽,盾甲龙,中龙,空尾蜥
代表化石:盘龙类(前期),兽孔类(后期),巨齿龙类,离片椎类,壳椎类,种子蕨,裸子植物(针叶类,苏铁类,银杏类)