

恐头兽亚目(Tapinocephalia )的祖先是外形类似巴莫鳄的中小型肉身动物,比如利齿狭头兽。它们的后代在中二叠世占据了非常多样的生态位。其中貘头兽类转为彻底的植食性,为了撸叶子甚至不惜退化掉了合弓纲祖传的异型齿。

这张图上回放了一半。上面六个头骨是安蒂欧兽类的,下面六个是貘头兽类(及其近亲)的。大略展示了植食性恐头兽是怎样失去锋利的犬齿的。图片来源自网络。
另一些恐头兽类保留甚至强化了牙齿的刺穿和撕咬功能。它们演化成了比貘头兽类更加巨大和怪异的杂食性和肉食性类群。井蛙觉得,“恐头”兽这个名字和它们更配。
杂食性恐头兽首推俄罗斯彼尔姆地区(Perm Region )出土的冠鳄兽( Estemmenosuchus )。因为头骨上伸出两对怪异的角,很是吸引眼球,所以三长两不短地总能在各种二叠纪古生物展上露个脸。

国外某博物馆的“二叠纪怪物展”模型。从前到后:巨型兽,锯齿龙,狼蜥兽,冠鳄兽,异齿龙。真是合弓纲出尽风头的时代。图片来源自[1]。
冠鳄兽属 Estemmenosuchus 包括两个种: Estemmenosuchus uralensis (乌拉尔冠鳄兽)和 Estemmenosuchus mirabilis (奇异冠鳄兽)。它们的区别在于:乌拉尔冠鳄兽体型更大,但是头上的冠饰(角)不明显;奇异冠鳄兽体型小得多,但是冠饰大,而且棱角分明,更加美观。许多复原形象为了照顾视觉效果,把两种冠鳄兽的优点杂糅到一起了。
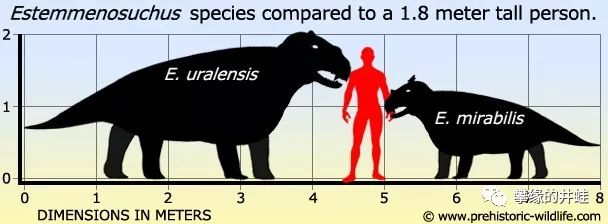
两种冠鳄兽的体型对比。显然乌拉尔冠鳄兽(左)的身姿要雄壮得多。图片来源见水印。

乌拉尔冠鳄兽的头骨化石和复原图。不过这个大个子的角和体型实在有点不搭配。图片来源自网络。

乌拉尔冠鳄兽骨架化石。不仔细看,根本注意不到头上的“角”。图片来源自网络。

奇异冠鳄兽的头骨化石。它的角就帅气多了。图片来源自Wikimedia。
看冠鳄兽的牙齿,它们显然应该更偏向肉食性,或者像现代野猪一样,有什么吃什么。水中的鱼虾水草,植物的嫩芽和根茎,动物的活体或者尸体,一概不挑。至于头上的“角”,人们大多认为是同类间展示或者争斗之用,功能类似今天牛羊的角或者河马的长牙。对此,井蛙有一些不同的猜测,后面再说。

一群奇异冠鳄兽的生态复原图。话说,用这样的獠牙啃草,总觉得不对劲。图片来源自网络。
冠鳄兽属于独立的冠鳄兽科(Estemmenosuchidae),从生存年代(大约2.67亿年前)来看,属于恐头兽类的一个早期分支。和冠鳄兽类似的杂食性恐头兽类还有戟头兽科(Styracocephalidae),多特罗龙兽科(Deuterosauridae)和伟鳄兽科(Titanosuchidae),这几个科曾经被归入貘头兽类或者安蒂欧兽类,但是比较新的文献似乎倾向于把它们独立出来[2]。

戟头兽Styracocephalus platyrhynchus头骨结构和复原图。它的生存年代(约2.6到2.65亿年前)略晚于冠鳄兽。化石于1929年在南非被发现[3]。图片来源自网络。
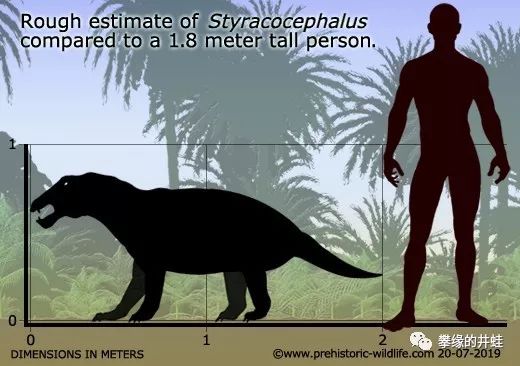
戟头兽体型对比。它还长有不大的犬齿,大概处在从肉身性像植食性转变的中间阶段。图片来源见水印。

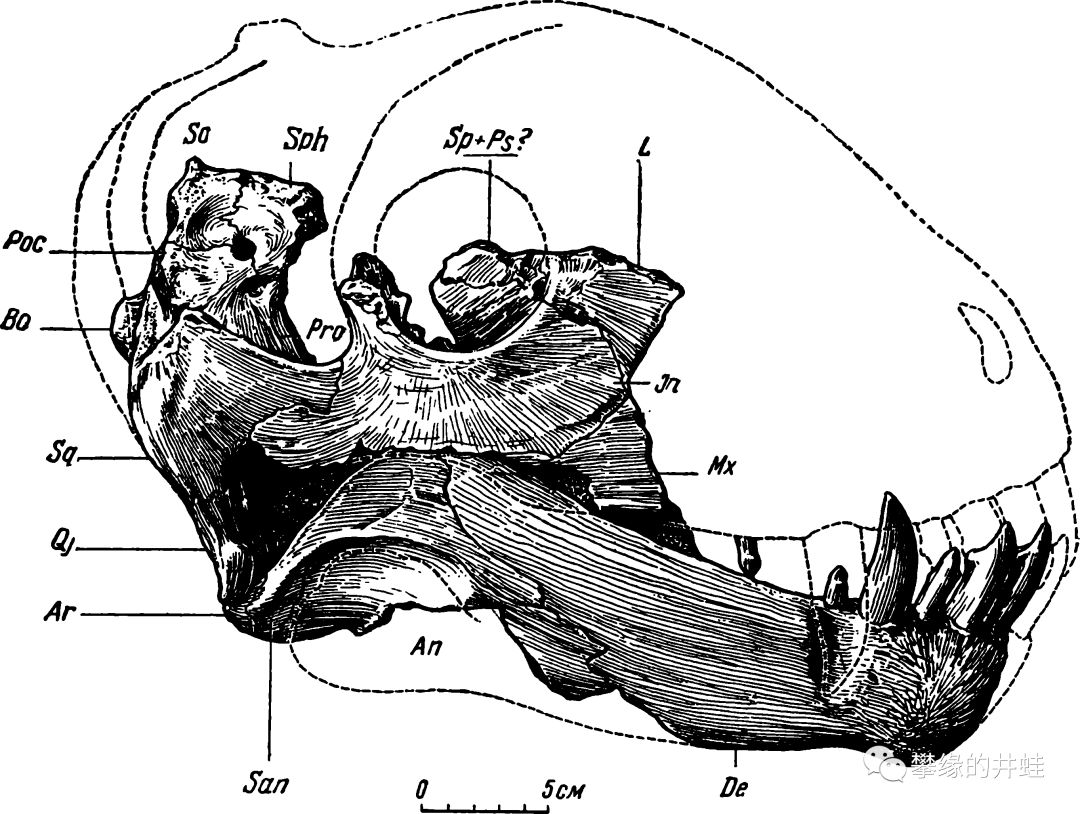
俄罗斯出土的多特罗龙兽Deuterosaurus biarmicus化石大多是头骨的碎片。有的头骨足有80厘米,而且特别厚重。推测这种动物全长可能有5到6米,而且极其粗壮。图片来源自网络。

多特罗龙兽复原图。它大概是中二叠世的熊吧。图片来源见水印。
伟鳄兽科里有两个大家伙:姜氏兽( Jonkeria )和伟鳄兽( Titanosuchus )。化石也都在南非发现(中二叠世的南非挤满了各种恐头兽啊)。姜氏兽1916 年定名,伟鳄兽更早,1876 年定名。但是这一百多年人们看各种中生代蜥形纲巨怪看花了眼,感觉这些南非的恐头兽自从发现后就被打入冷宫,很少有人研究发新论文,网上的资料更是少得可怜。

姜氏兽头骨化石。图片来源自网络。

一群姜氏兽复原图。有人认为姜氏兽吃草,有人认为吃肉,我觉得可能和冠鳄兽一样,什么都吃。图片来源见水印。
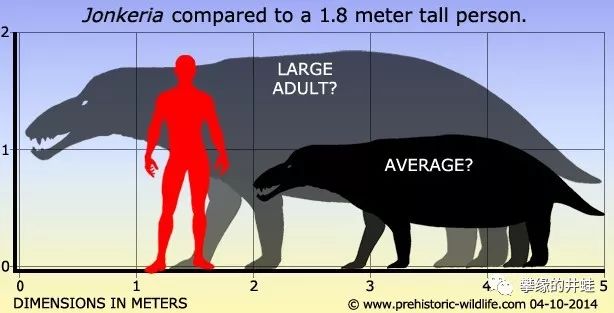
姜氏兽(可能)的体型。这个属(Jonkeria)的分类很乱,感觉好像每找到一块化石,发现者就赶紧跑去命名一个新种。光是文献里出现过的就有:J.truculenta,J. boonstrai,J.haughtoni,J. ingens,J.koupensis,J. parva,J.rossouwi,J. vanderbyli,想想就知道肯定有好多重复的。这么多年了也没人系统地整理一下,果然冷门。图片来源见水印。

在网上,伟鳄兽连张化石照片都找不到。井蛙只知道这是一种大型食肉兽,体长大约2.5米,可能以同时期的麝足兽和姜氏兽为食。上图是伟鳄兽猎捕麝足兽的想象图,来源见水印。
一般认为上面这些恐头兽是杂食性的种类,不过井蛙觉得它们应该还是以肉食为主——我不相信在二叠纪有那么多熊猫。恐头兽类的另一大分支:安蒂欧兽类(Anteosauria )则是确定无疑的掠食者。它们盘踞在中二叠世食物链的顶端。
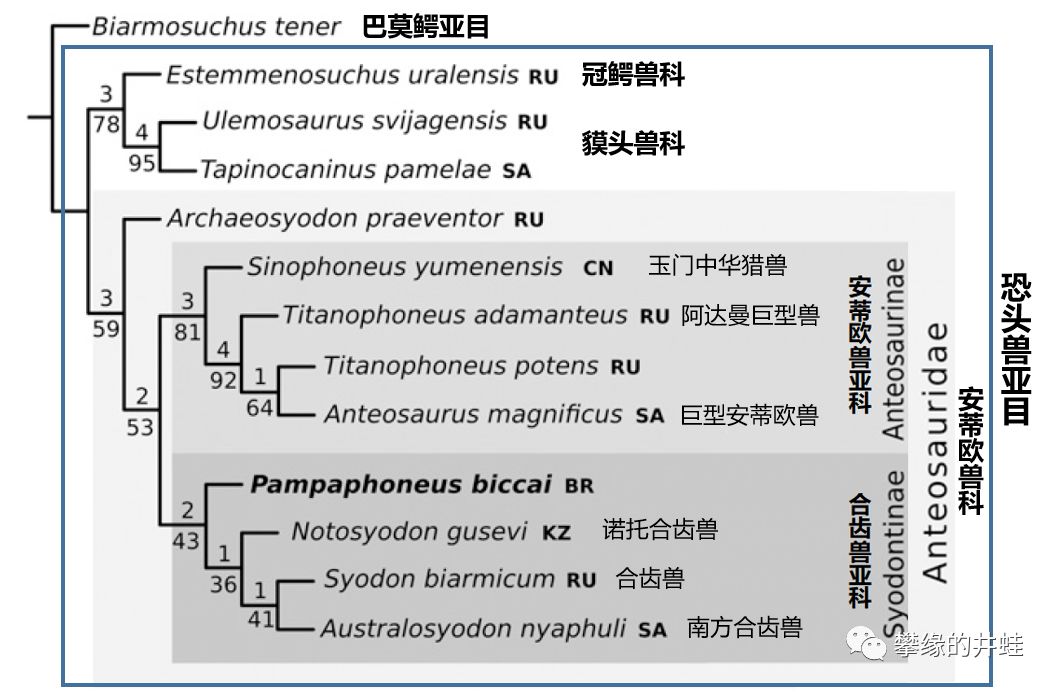
安蒂欧兽科下右两个亚科:安蒂欧兽亚科Anteosaurinae和合齿兽亚科Syodontinae。相对而言,安蒂欧兽亚科成员体型大而粗壮,合齿兽亚科成员体型小而苗条。二者的地位大概类似于中二叠世的狮虎和豺狼吧。图片来源自[4]。

安蒂欧兽亚科核心成员的集体复原图。左前是著名的巨型安蒂欧兽(Anteosaurus magnificus),右前是我国发现的玉门中华猎兽(参见上回),后面三只是巨型兽属的成员。图片来源自网络。

合齿兽亚科核心成员的集体复原图。这个科中的属名相当整齐划一:合齿兽属(Syodon),诺托合齿兽属(Notosyodon),小合齿兽属(Microsyodon),古合齿兽属(Archaeosyodon),南方合齿兽属(Australosyodon)。唯一的例外是2008年在巴西找到的新属种Pampaphoneus biccai。图片来源自网络。

中二叠世安蒂欧兽亚科和合齿兽亚科的分布。对应今天的化石发现地:1,俄罗斯平原;2,乌拉尔山南麓;3,中国西部;4,5,巴西巴拉那盆地;6,南非卡鲁盆地。这些游荡在返古大陆上的掠食者是所有其他动物的噩梦。图片来源自[4]。
中二叠世最强大的超级掠食者首推南非的安蒂欧兽属( Anteosaurus ,上图中最下面的灰色图标)和俄罗斯的巨型兽属( Titanophoneus ,上图中上方的两个灰色图标)。它们都属于安蒂欧兽亚科,虽然一南一北相距遥远,却有着很近的亲缘关系。
已发现的最大的安蒂欧兽头骨化石 Anteosaurus magnificus 长达80 厘米,这只动物活着的时候也是5 ,6 米长,足以猎杀麝足兽的怪物。


巨型安蒂欧兽头骨化石和全身比例图。图片来源自网络。
巨型兽属有两个大型种:阿达曼巨型兽 Titanophoneus admanteus 和博滕斯巨型兽 Titanophoneus potens 。关于这两类巨型兽的体型,各种资料提供的数据并不统一。井蛙看到的最大记录是:

博滕斯巨型兽,头骨全长65厘米左右。图片来源自网络。

阿达曼巨型兽,头骨全长80厘米左右。图片来源自网络。
基本上,它们的体型都和巨型安蒂欧兽有一拼。

一些比较小(可能是幼年期)的博滕斯巨型兽留下了比较完整的骨架。巨型安蒂欧兽和阿达曼巨型兽的全身复原基本上都是参考它来的。图片来源自网络。
合齿兽亚科的体型比安蒂欧兽亚科要小上几号,体长大约在1 到3 米左右,占据着中型掠食者的生态位。也许就是它们把巴莫鳄类逼成了小巧的伯纳特兽类(参见第二百四十回百兽图4 :盖凿齿)。
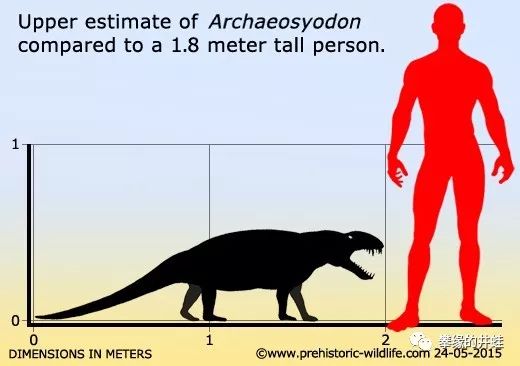

古合齿兽(Archaeosyodon)和南方合齿兽(Australosyodon)的体型。感觉好像也不是打不过的样子。除非,它们是成群狩猎的。图片来源见水印。
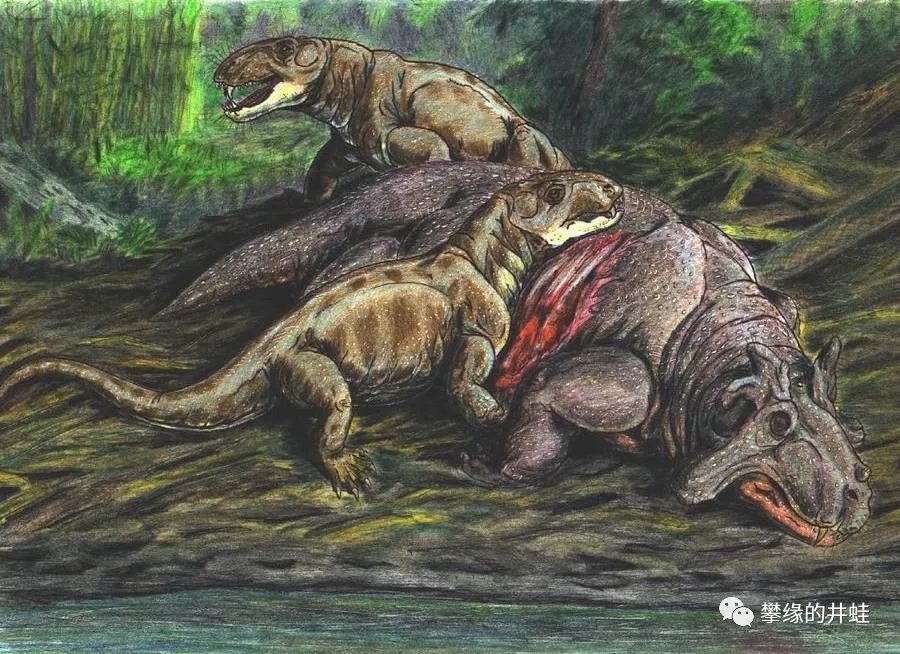
上图是想象中的古合齿兽群猎杀冠鳄兽。老实说,井蛙认为即使是小个子的奇异冠鳄兽成体,也不是一群南方合齿兽能干掉的,说是找到了现成的尸体正在食腐还差不多。图片来源自网络。

俄罗斯博物馆里的合齿兽Syodon efremovi头骨化石。和安蒂欧兽,巨型兽一比,秀气不少啊。图片来源自网络。
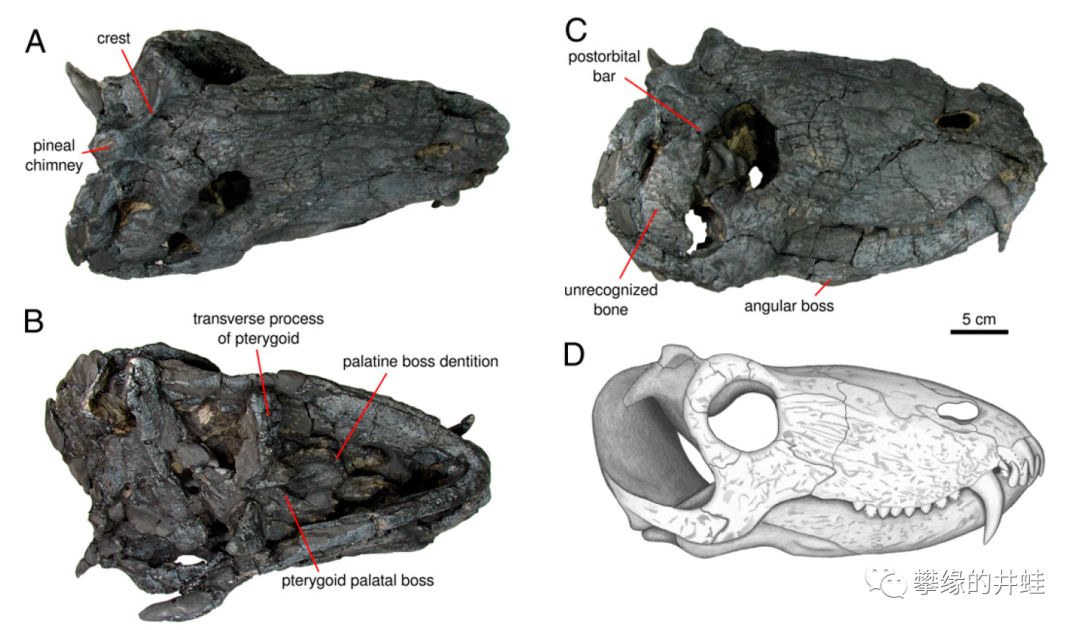
巴西巴拉那盆地(就是上面地理分布图中5那个位置)新发现的合齿兽类Pampaphoneus biccai头骨化石和复原图。全长3米左右,在合齿兽中算是最大的了。它的发现勾勒出了安蒂欧兽科在返古大陆上的迁移路径(地理分布图中的红色箭头)。图片来源自[4]。
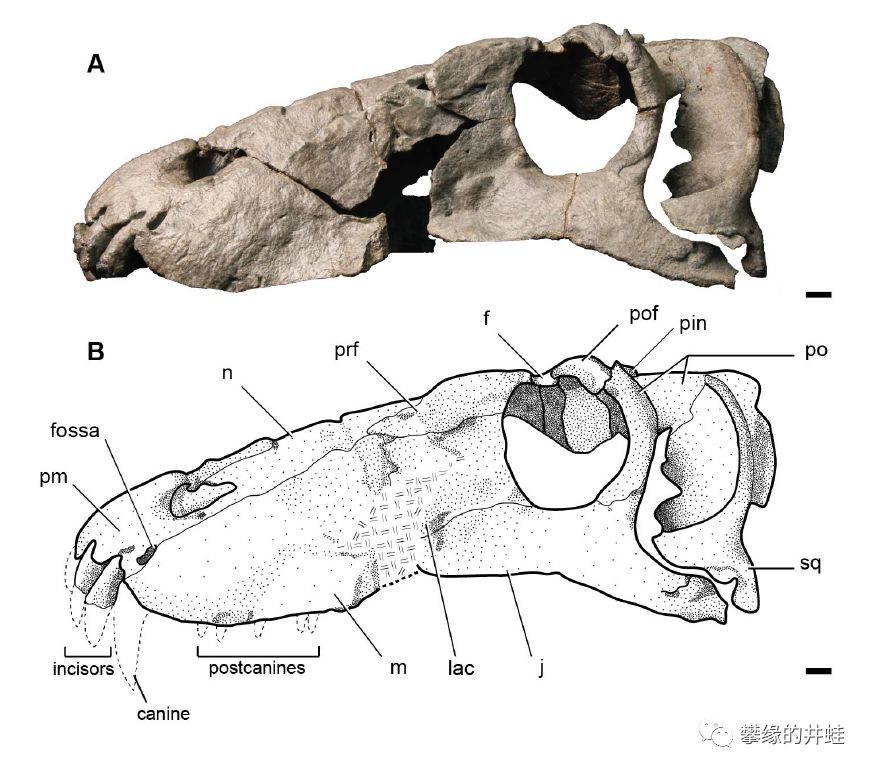
这块头骨标本来自南非卡鲁盆地,编号BP/1/7074,长28厘米。看起来很像合齿兽,但研究者认为它是一头幼年的巨型安蒂欧兽。图片来源自[5],标尺长度1厘米。
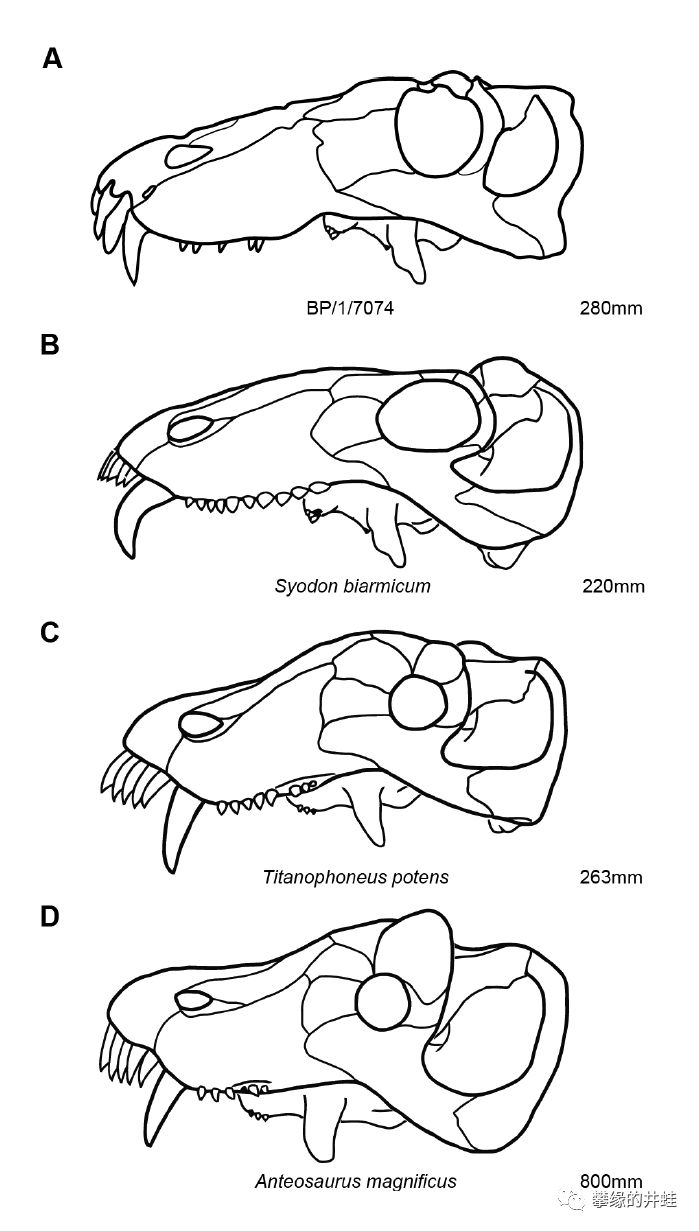
BP/1/7074号头骨(A),合齿兽(B),博滕斯巨型兽(C),巨型安蒂欧兽成体头骨(D)的结构对比。看起来,巨型兽和安蒂欧兽就像是合齿兽的强化版。图片来源自[5]。
不知大家发现没有,所有介绍过的恐头兽类中,体型最大的不是植食性的貘头兽类(比如麝足兽),而是肉食性(安蒂欧兽和巨型兽)和杂食性(冠鳄兽,姜氏兽和多特罗龙兽)的种类,取个学名又是Titano- 又是Meganifi- 的。而在中生代和新生代的陆地生态系统中,大型植食动物的体型要远远超过大型肉食动物。
这种二叠纪特有的体型倒挂也许反映了两个问题。一是貘头兽类的消化能力堪忧。它们没有演化出诸如胃石,反刍或者盲肠这样的辅助手段,囫囵吞下的叶片通过肠胃,只能榨出很少的一点营养,供养不起太过庞大的身体。而从动物性食物中摄取蛋白质和脂肪就方便多了,也更容易长出巨大的体型。
另一个原因可能出在肉食(杂食)恐头兽身上:它们没有高效击杀大型猎物的手段。犬齿不够长,咬合力不够大,不能高速奔跑,也没有锋利的爪子,逼得它们必须依靠更大的躯体和力量去制服猎物。井蛙怀疑,冠鳄兽那奇怪的头饰,很可能是用来攻击猎物的。仔细看安蒂欧兽和巨型兽的头骨,上面也有肿大的瘤块。它们也许会用巨头把猎物撞翻,然后踩住撕咬。

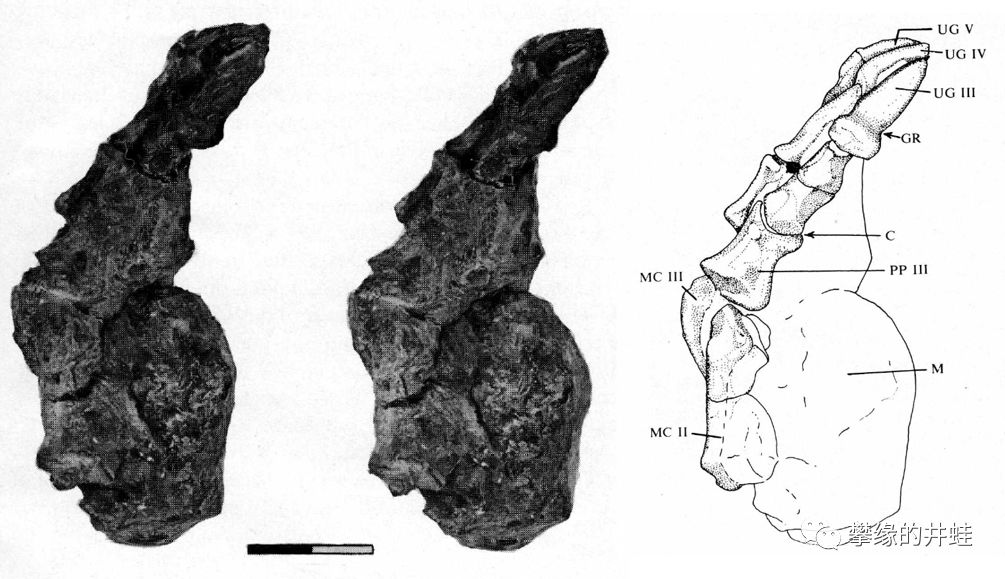
这是一只安蒂欧兽Micranteosaurus parvus(很可能就是一只幼年的巨型安蒂欧兽)的脚掌化石。实在看不出有多强的杀伤力啊,捕食的时候,与其说用利爪撕碎猎物,更像是大嘴巴抽晕猎物吧。图片来源自[6],标尺长度2厘米。
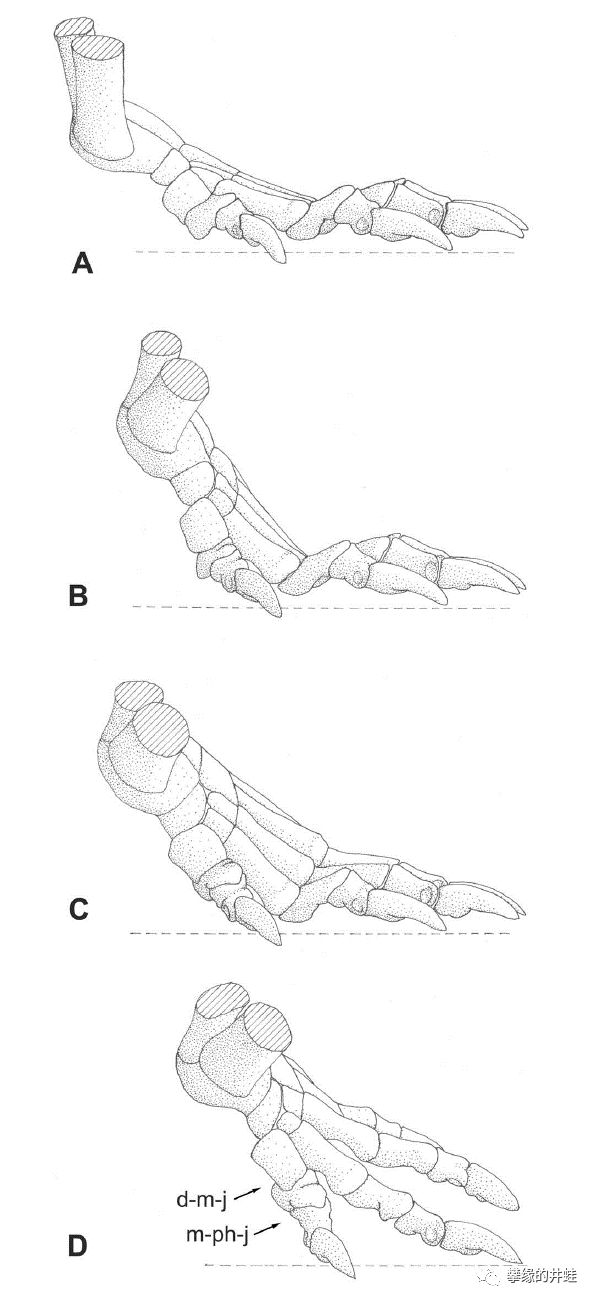
巨型兽爬行时前脚掌的支撑方式。这小内八撇的,能跑多快?大概反正它的猎物也都是这么爬的,速度应该够用。图片来源自[7]。
也许正是因为这种种缺陷,导致恐头兽类在中二叠世末期遭到全灭。锯齿龙和大型二齿兽瓜分了貘头兽类的生态位;而安蒂欧兽,巨型兽和合齿兽,则被丽齿兽亚目(Gorgonopsia )的新型掠食者取代。“Gorgonopsia ”源自希腊神话里的蛇发女妖戈耳工(Gorgon ),我猜我国的前辈古生物学家感受到了“蛇发女妖兽”这个名字扑面而来的中二气息,所以译成了“丽齿兽”。信达雅,佩服!
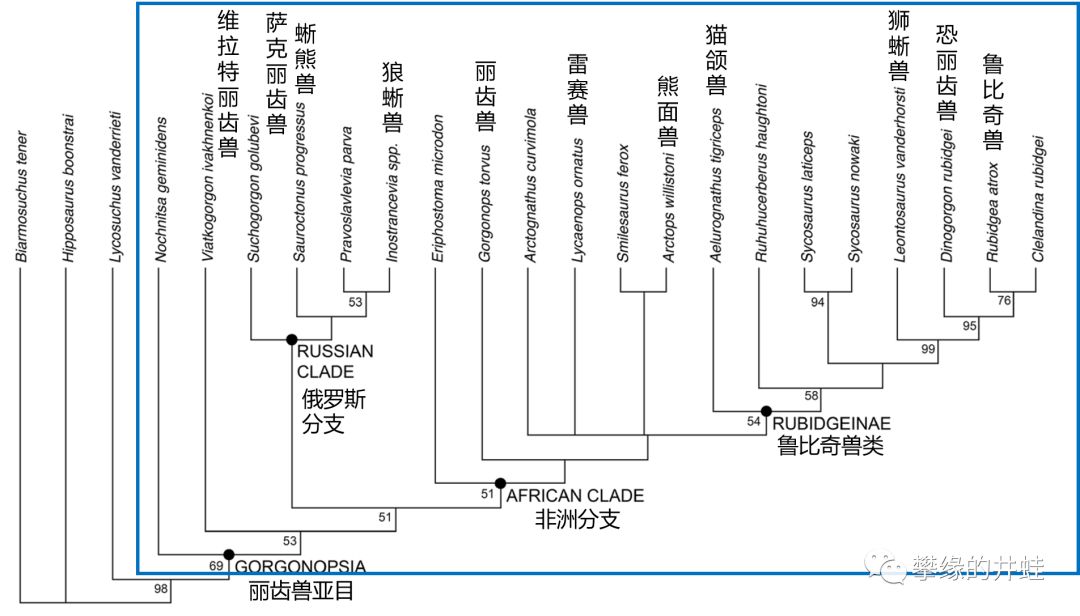
丽齿兽亚目演化谱系和主要种类。大家可以试着把厘米的“丽齿兽”都换成“蛇发女妖兽”,中二度直接爆表。右数第七个“Ruhuhucerberus haughtoni”直译是“鲁胡胡地狱三头犬兽”,井蛙也想试着翻个信达雅的译名,奈何功力不够,做不到啊(大家觉得“酆都兽”怎么样?)。图片来源自[8]。
从已发现的化石来看,丽齿兽类的身体结构要全面优于肉食性恐头兽类。它们是更加高效的杀戮机器。
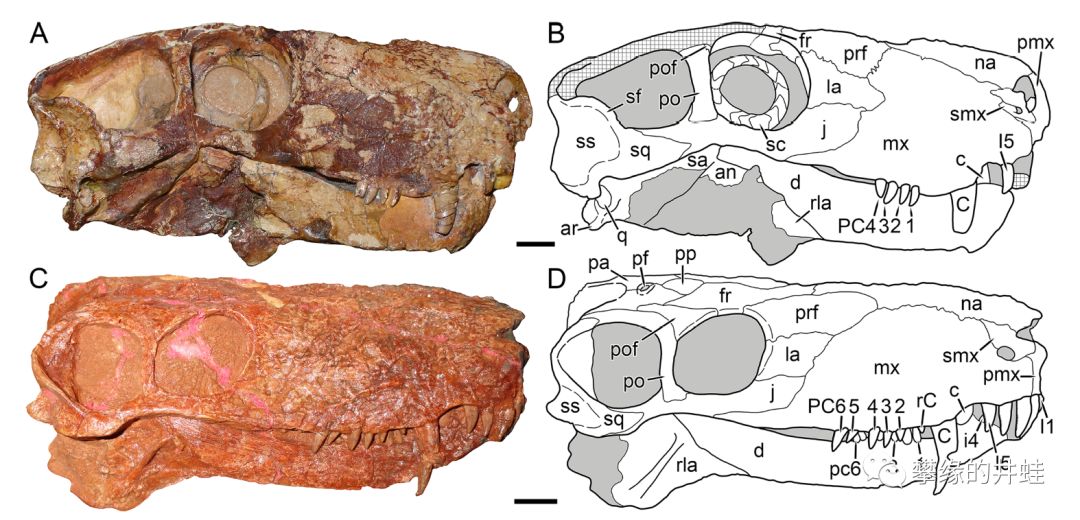
俄罗斯发现的两种丽齿兽头骨化石。A,B:维拉特丽齿兽Viatkogorgon ivakhnenkoi;C,D:Nochnitsa geminidens。与肉食性恐头兽相比,丽齿兽类拥有更发达的犬齿,牙齿的生长角度和上下颌比例更合理(安蒂欧兽和合齿兽是典型的地包天,这种下颌能提供的咬合力很有限)。图片来源自[8],标尺长度1厘米。
丽齿兽类脑后那对大得夸张的颞颥孔可以容纳异常粗大的肌肉,配合壮硕的下颌,爆发出恐怖的咬合力。下面是一些鲁比奇兽类(丽齿兽亚目下的一个超科Rubidgeinae ,化石主要分布在南非)的头骨化石,感受一下它们的煞气吧(图片全部来自[9] ,标尺长度全部为10 厘米):

Clelandina rubidgei,一种小型的鲁比奇兽。

Smilesaurus ferox,比Clelandina rubidgei稍大一点。

猫颌兽Aelurognathus tigriceps 。

恐丽齿兽Dinogorgonrubidgei。

狮蜥兽Leontosaurusvanderhorsti。

鲁比奇兽Rubidgea atrox。

“鲁胡胡地狱三头犬兽”Ruhuhucerberus haughtoni。

Sycosaurus laticeps。

Sycosaurus nowaki。
这些丽齿兽的身体大小,可以参照下面这张图:

熊面兽Arctops willistoni头骨和身体的比例。这么看来,头骨长度到30厘米以上的,就已经是很恐怖的掠食动物了。图片来源自网络。
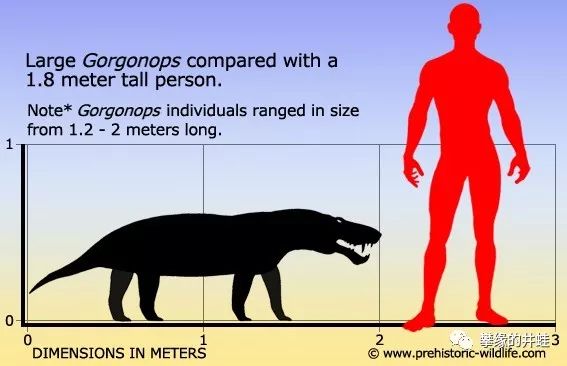
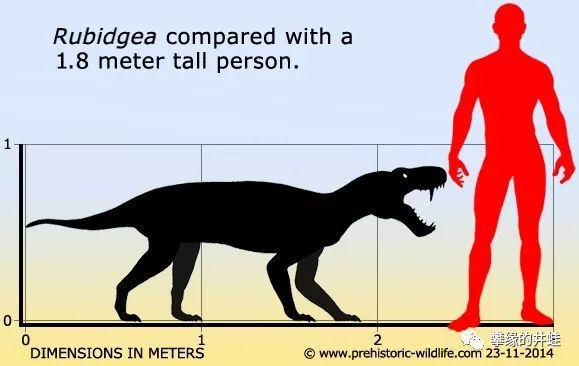

两米以上的大型丽齿兽类基本上就能轻松干掉一个成年人。图片来源见水印。
丽齿兽类的前后肢长度接近,支撑方式更接近直立姿态,比起恐头兽类轧悠轧悠的小内八,奔跑起来更加轻快省力。它们的尾巴也大为缩短,持平衡的同时减少了阻力。总之,这是一类更加迅捷矫健的掠食者。

雷赛兽Lycaenops ornatus骨架。和前面博滕斯巨型兽的骨架对比一下,显然丽齿兽类的体型更加紧凑,更像我们熟悉的“兽”了。图片来源自Wikimedia。
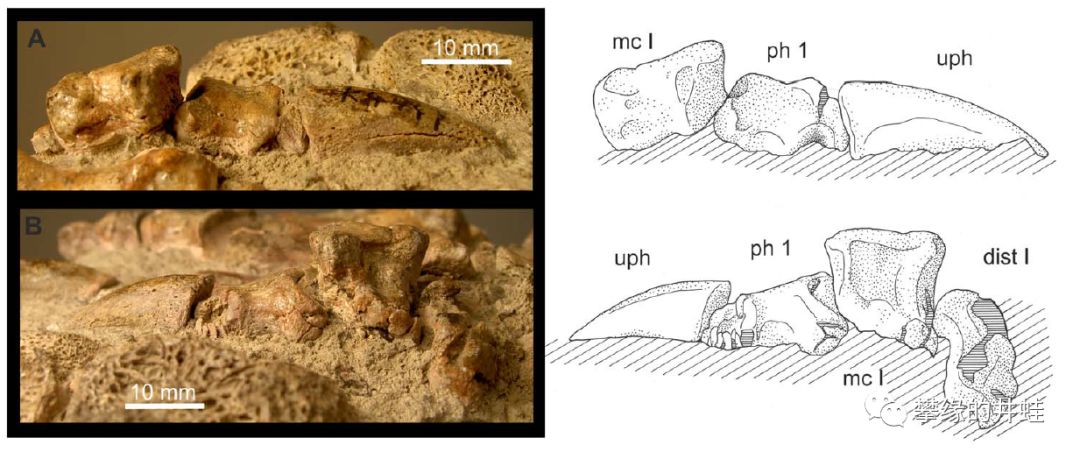
某种丽齿兽类的爪子。比起安蒂欧兽的大巴掌升级了不少。图片来源自[7]。
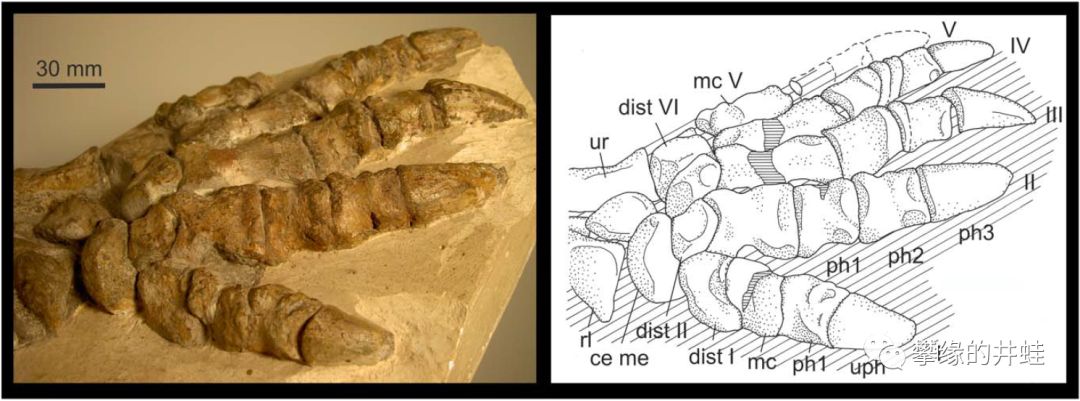
某种鲁比奇兽的足部化石,整只脚掌平按在地上。丽齿兽类的奔跑姿势可能像现代哺乳动物一样,弯腰收腹,前后肢交替。图片来源自[7]。
把上面这些致命武器装备齐全,再加上巨大的身体,终于早就了二叠纪最强大的肉食合弓兽:狼蜥兽。

博物馆陈列的大型狼蜥兽Inostrancevia alexandri骨架。图片来源自网络。
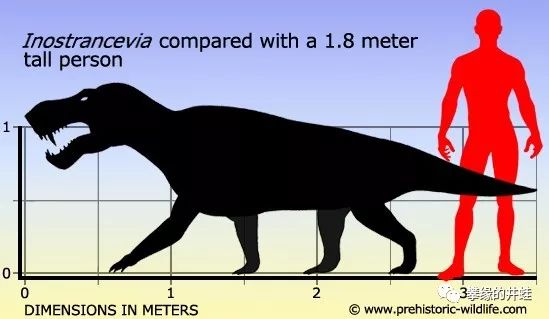
大型狼蜥兽的体型。它的体型比不上安蒂欧兽和巨型兽,但杀伤力绝不弱于恐头兽前辈。图片来源见水印。
狼蜥兽的身体结构几乎就是为了猎杀锯齿龙这样的重甲发酵罐设计的。巨大的身体的高速扑击,再加上锋利的剑齿和强大的咬合力,可以以最小的代价杀死并肢解猎物。井蛙猜测,大型丽齿兽类可能是第一批能够咬碎猎物骨骼的陆生食肉动物。

巨大的肉食合弓兽是二叠纪的霸主,更是合弓帝国的象征。它们磨牙吮血,所向披靡。图片来源自网络。

然而命运开了个天大的玩笑,到了三叠纪,蜥形纲和合弓纲的地位就彻底翻转了。图片来源阻网络。
丽齿兽亚目属于兽孔目中的“兽齿类(Theriodontia )”,这个分类单元还包括兽头亚目(Therocephalia )和犬齿兽亚目(Cynodontia ),后两者属于演化程度更高的“真兽齿类(Eutheriodontia )”。当二叠纪的无限风光成了一场幻梦,丽齿兽亚目随着合弓王朝的倾覆灰飞烟灭,兽头亚目和犬齿兽亚目则在中生代惨淡求生,等待不知何时命运的玩笑再一次降临。它们的故事,就留待下回分解吧。

兽齿类三个亚目的头骨对比。1:恐丽齿兽(丽齿兽亚目);2:麝喙兽Moschorhinus(兽头亚目);3:犬颌兽Cynognathus(犬齿兽亚目)。哺乳动物就处在这个演化支的最末端。图片来源自[10],标尺长度10厘米。
地球名片
生物分类:动物界-脊索动物门-脊椎动物亚门-羊膜类-合弓纲-兽孔目-丽齿兽亚目
存在时间:二叠纪中期至二叠纪末
现存种类:无
化石种类:未找到完整统计
生活环境:陆地
代表特征:更接近直立步态,更紧凑的身体,发达的剑齿,巨大的颞颥孔
主要种类:丽齿兽,恐丽齿兽,鲁比奇兽,狼蜥兽,熊面兽,雷赛兽,“鲁胡胡地狱三头犬兽”
参考文献
[1] https://gondwanastudios.com/gallery/
[2] Saniye Atayman, Bruce S. Rubidge, Fernando Abdala,Taxonomic re-evaluation of tapinocephalid dinocephalians. Research, 2016, DOI:10.13140/RG.2.1.3932.6482
[3] S. H. Haughton, On some new therapsid genera. Annals ofthe South African Museum 28:55-78, 1929
[4] Juan Carlos Cisneros, Fernando Abdala, SaniyeAtayman-Güven, et al., Carnivorous dinocephalian from the Middle Permian ofBrazil and tetrapod dispersal in Pangaea. PNAS, vol. 109, 1584–1588, 2012
[5] Ashley Kruger, Ontogeny and cranial morphology of thebasal carnivorous dinocephalian, anteosaurus magnificus from the tapinocephalusassembage zone of the South African Karoo. A Dissertation submitted to theFaculty of Science, University of the Witwatersrand, Johannesburg, infulfilment of the requirements for the degree of Master of Science, 2014
[6] T. Rowe, J. A. van den Heever, The hand of Anteosaurusmagnificus (Dinocephalia: Therapsida) and its bearing on the origin of themammalian manual phalangeal formula. Suid-Afrikaanse Tydskrif vir Wetenskap,Vol. 82, 641-645, 1986
[7] Kümmell SB, Frey E, Range of Movement in Ray I of Manus andPes and the Prehensility of the Autopodia in the Early Permian to LateCretaceous Non-Anomodont Synapsida. PLoS ONE 9(12): e113911, doi: 10.1371/journal.pone.0113911,2014
[8] Kammerer and Masyutin (2018), Gorgonopsian therapsids(Nochnitsa gen. nov. and Viatkogorgon) from the Permian Kotelnich locality ofRussia. PeerJ 6:e4954; DOI 10.7717/peerj.4954
[9] Kammerer, Systematics of the Rubidgeinae (Therapsida:Gorgonopsia). PeerJ 4: e1608, DOI: 10.7717/peerj.1608, 2016
[10] BLAIRE VAN VALKENBURGH, IAN JENKINS, EVOLUTIONARYPATTERNS IN THE HISTORY OF PERMO-TRIASSIC AND CENOZOIC SYNAPSID PREDATORS. PALEONTOLOGICALSOCIETY PAPERS, V. 8, 267-288, 2002