

二叠纪篇终于进入最高潮:合弓纲的狰狞怪兽们登场了。
古老的盘龙目(参见第二百回百兽图1:帆影;第二百一回百兽图2:大脚;第二百二回百兽图3:鸭子)筚路蓝缕,拓土开疆,奠定了陆地霸主的基业,然后把这大好江山交给了演化程度更高的类群。

盘龙目各主要类群在石炭纪晚期到二叠纪的多样性变化情况。挺着背帆的基龙和异齿龙,大脚草包杯鼻龙,还有像蜥蜴的蜥代龙和像鳄鱼的蛇齿龙,全都在二叠纪中期黯然退场。图片来源自[1]。

盘龙目(Pelycosauria)代表着合弓纲尚处于“蜥蜴形”的原始阶段。已知生存时间最晚的盘龙类是生活在二叠纪中期卡鲁盆地的一种蜥代龙。它的退场,标准着合弓纲彻底弃守“蜥蜴形”的基本构型,向着全新的演化阶段迈进。图片来源自网络。
在二叠纪中后期称霸陆地的合弓类被称为“兽孔目(Therapsida)”。这个演化支目前承盘龙目,后启哺乳类。它们的身体结构也正好介于二者之间,似龙非龙,似兽非兽的形态很容易让人联想起《山海经》里的种种神异怪物。
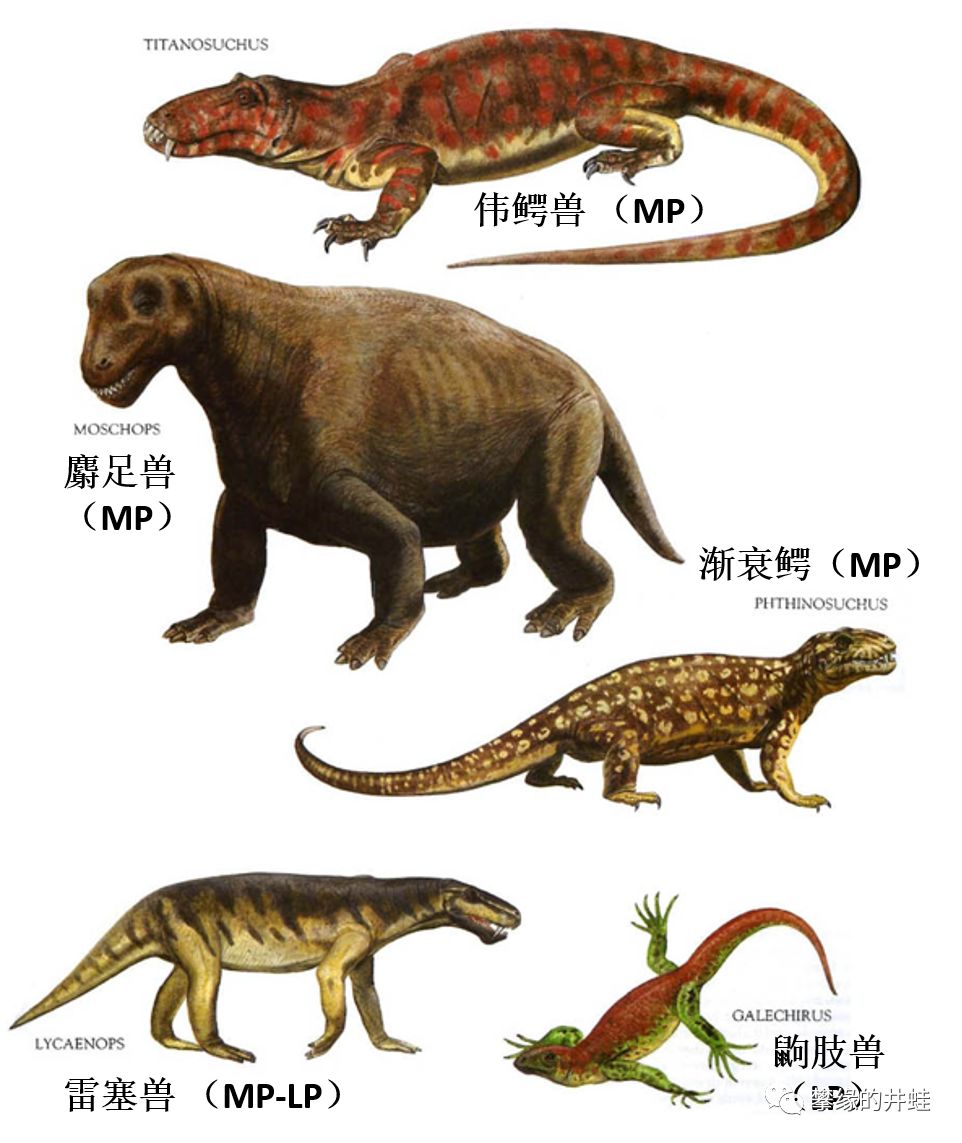
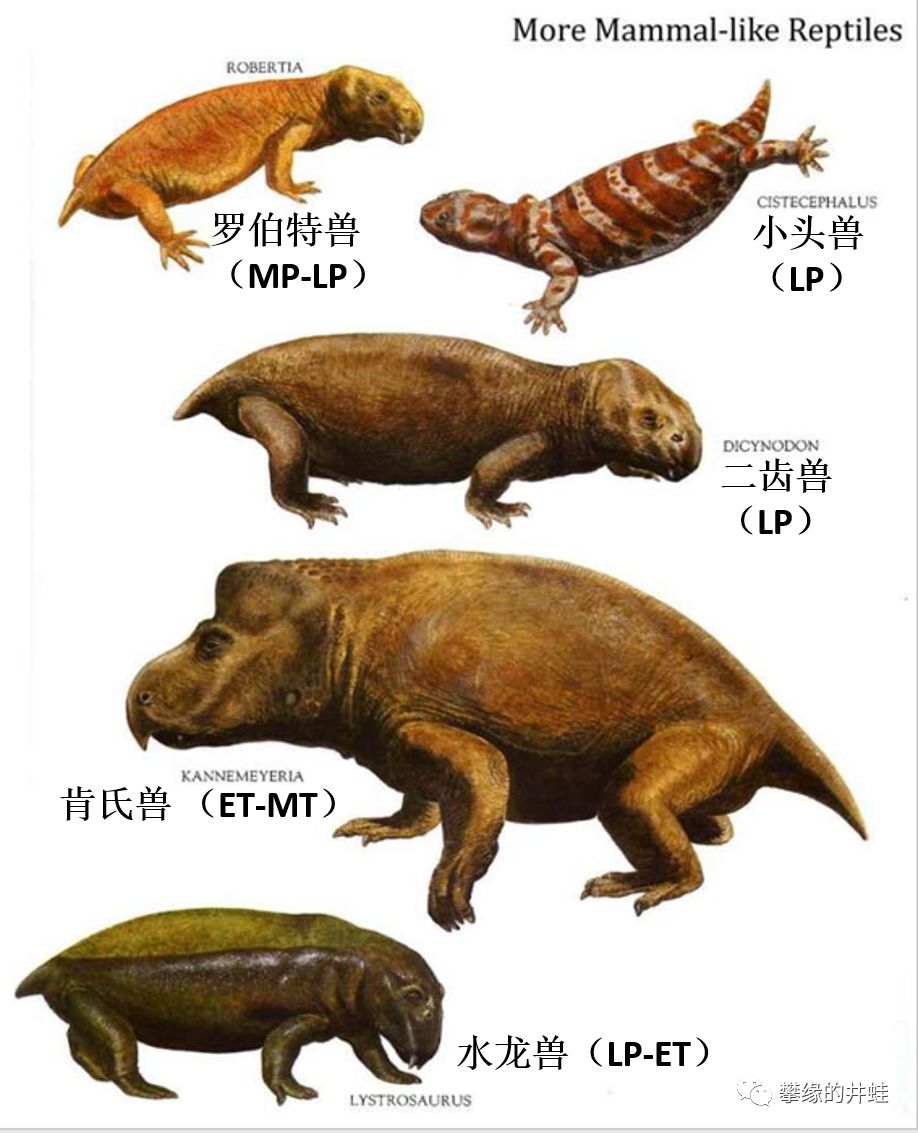

一些兽孔目动物的复原图,井蛙加上了译名和生存年代(MP:中二叠世;LP:晚二叠世;ET:早三叠世;MT:中三叠世;LT:晚三叠世;EJ:早侏罗世)。兽孔目中不乏比这些长相更加怪异的成员。图片来源自网络。
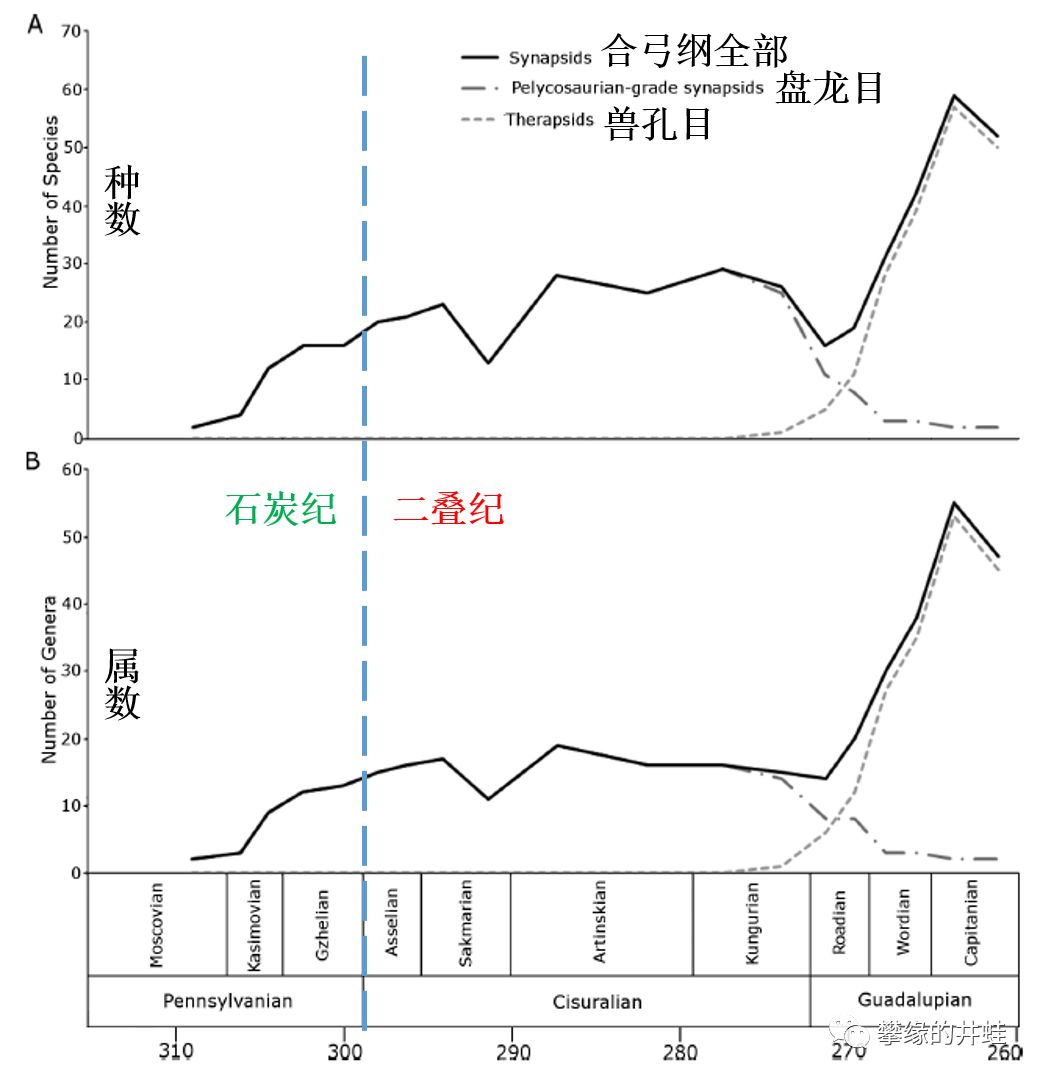
盘龙目和兽孔目多样性变化对比。可以看到大约在2.75亿年前,盘龙目跌落的同时,兽孔目迅速崛起,把合弓纲的多样性推上了一个高峰。图片来源自[1]。
除了种类和数量的激增,兽孔目的分布范围也比盘龙目广大得多,它们把合弓帝国的疆域推进到泛古大陆的各个角落。这也足以说明它们拥有更加强悍的生存能力。
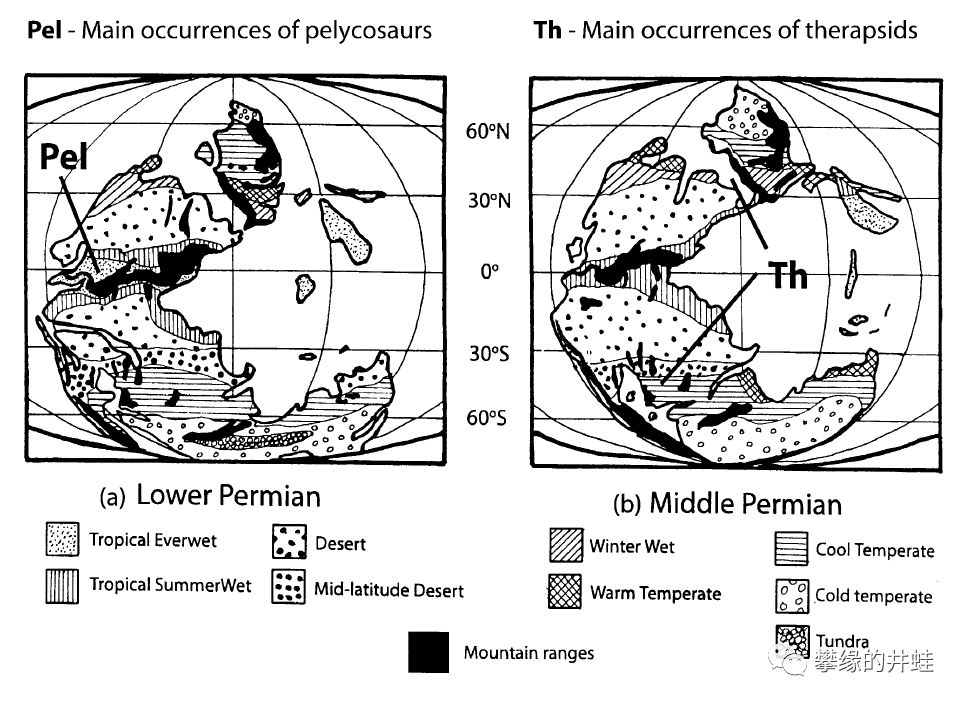
盘龙目(A图,Pel)和兽孔目(B图,Th)主要化石发现地区的二叠纪古气候对比。可以看到盘龙目主要分布在温暖湿润的热带雨林和湿地;而兽孔目已经扩散到原理赤道,寒冷干燥的内陆地区。图片来源自[2]。
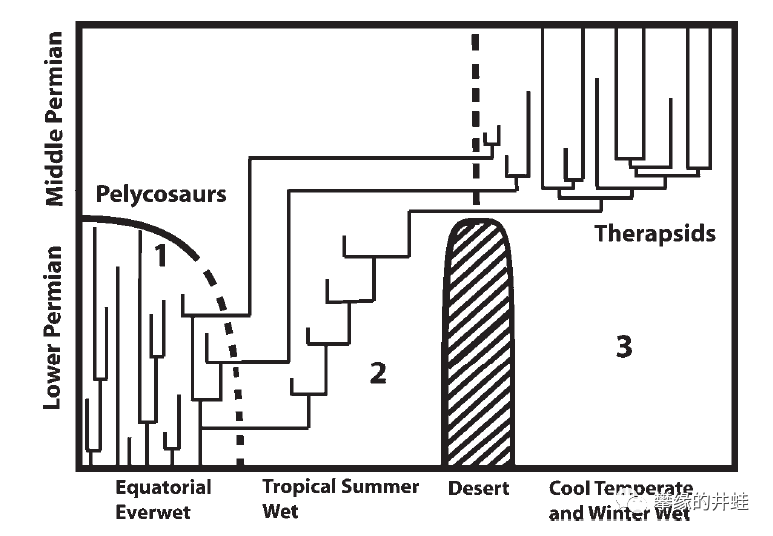
盘龙目和兽孔目的生存年代和分布范围示意图。显然,兽孔目比它的前辈更加适应干旱和寒冷的气候。由于缺乏软体组织的话说资料,我们并不能确定兽孔目究竟获得了怎样的装备,也许它们的羊膜卵结构更复杂,更加保温保水;也许是皮肤更加坚韧,甚至有毛发覆盖;也许代谢水平更高可以产生更多热量,甚至在某种程度上可以保持体温恒定。图片来源自[3]。
人们了解兽孔目动物,一靠化石发现,二靠和现存哺乳动物对比。但是哺乳动物只是一小撮兽孔类的直系后代,和大多数兽孔目动物的差异太大,而化石证据又太过残缺。我们至今还不能确定兽孔目起源的准确时间,以及最早的兽孔类到底长什么样子。
目前公认的最早的兽孔目动物是德克萨斯州发现的二叠纪早期的四角兽Tetraceratops insignis。只发现了一块严重扭曲变形的头骨。

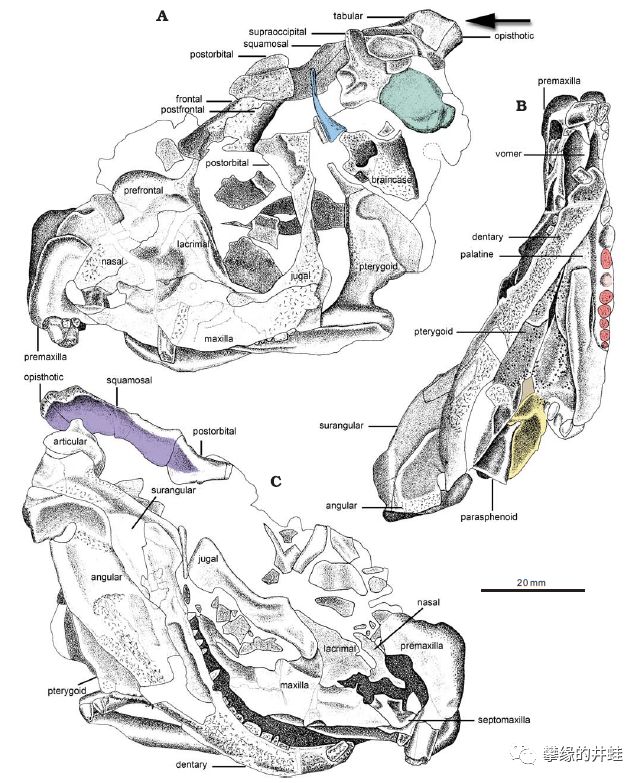
四角兽的头骨化石。基本上,大部分兽孔目动物都只留下完整或者残破的头骨,也许是因为它们的脑袋比较硬吧。图片来源自[4]。
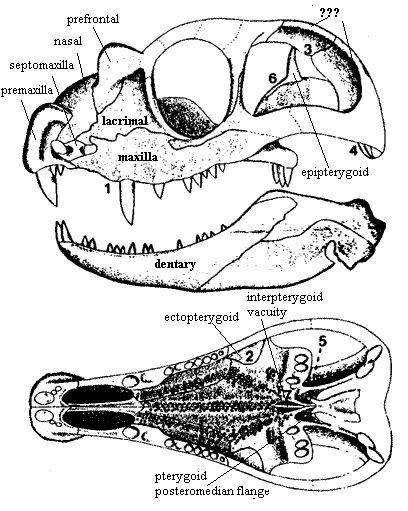
四角兽头骨复原图。它的额头和鼻尖有四个骨质突起,所以得名“四角兽”。图片来源自网络。

四角兽复原图。因为只发现了头骨,无从知道身体的样子,画成这样还是比较严谨的。图片来源自Wikimedia。
比四角兽晚一些的,有甘肃玉门大山口动物群发现的大山口珍稀兽Raranimus dashankouensis。不过它的化石就更残缺了,只有鼻骨和上颌。
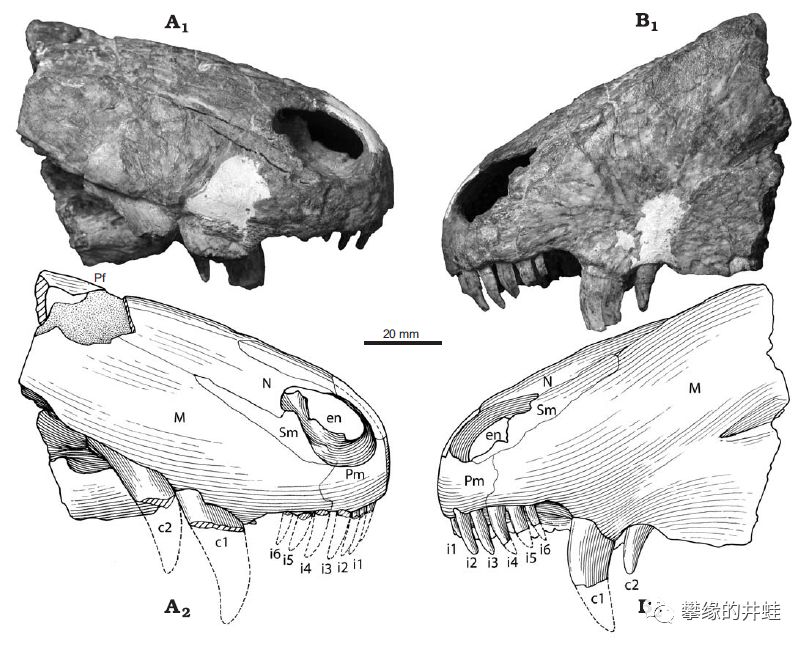
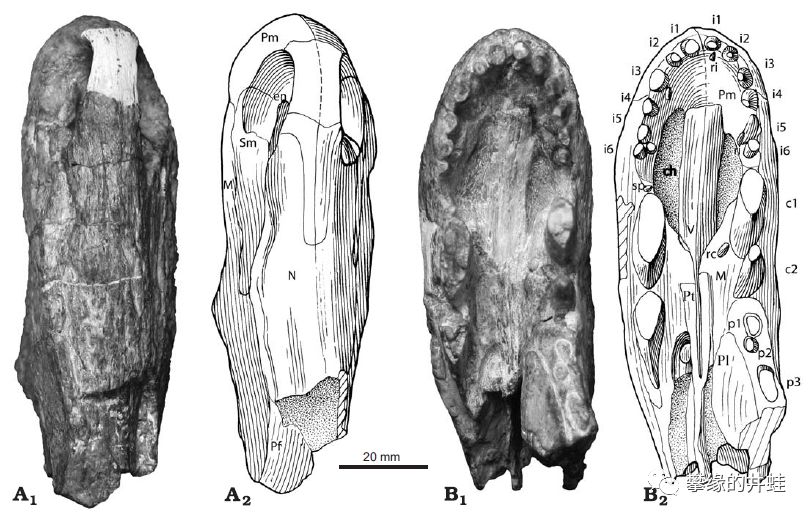
大山口珍稀兽化石。尽管化石资料如此缺少,但看到那两对特别显眼的“虎牙”,就可以确定它属于兽孔目。图片来源自[5]。
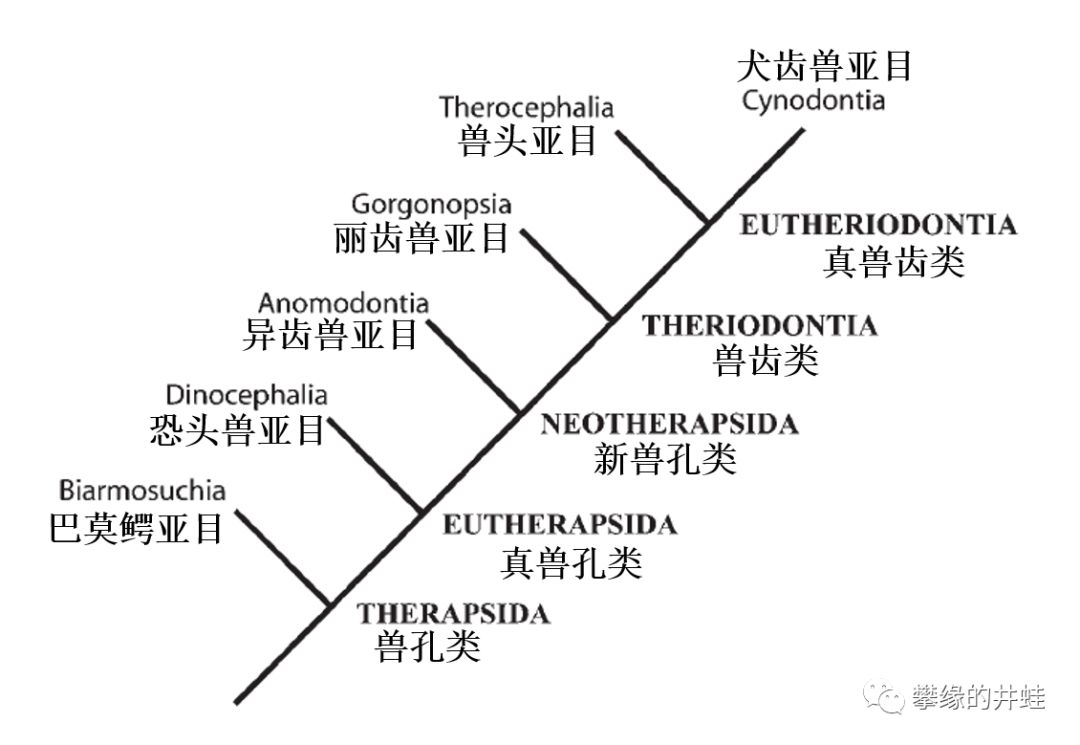
兽孔目主要类群的演化关系。一眼看去,除了“兽”之外,重复最多字的还有“头”“孔”和“齿”,颅骨,颞颥孔和牙齿固然是分类的重要依据,也是因为兽孔目留下的化石主要以头部居多的缘故。图片来源自[3]。
四角兽和其他早期兽孔类的头骨结构非常接近盘龙目的楔齿龙类,二者有很近的亲缘关系,也有人认为兽孔目正是起源于某种原始的楔齿龙类。
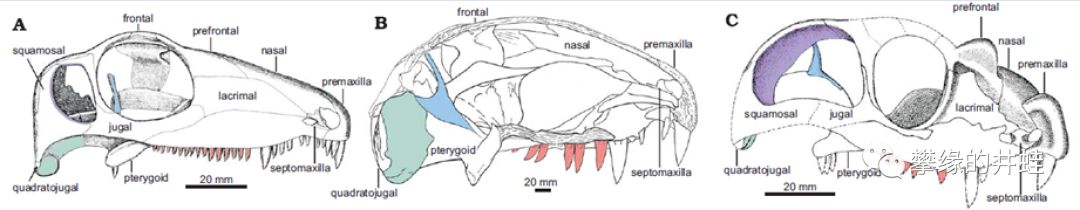
A:哈普托兽Haptodusgarnettensis(一种生活在晚石炭世到早二叠世的早期楔齿龙类);B:异齿龙Dimetrodon limbatus(头骨纵截面);C:四角兽的头骨结构对比。不难看出它们的相似之处。图片来源自[4]。
楔齿龙的牙齿已经开始出现分化,早期兽孔目显然更进一步,许多类群都演化出一对特别显眼的“剑齿”。这是非常强大的生存武器。食肉动物可以用它击破猎物的护甲,撕裂肌肉,刺穿血管和内脏,迅速解决战斗,节省体力的同时,也减少了猎物挣扎反击造成的伤害。植食和杂食的种类也可以用獠牙挖掘植物的根系,自卫反击,或者用于同类间的争斗和炫耀。
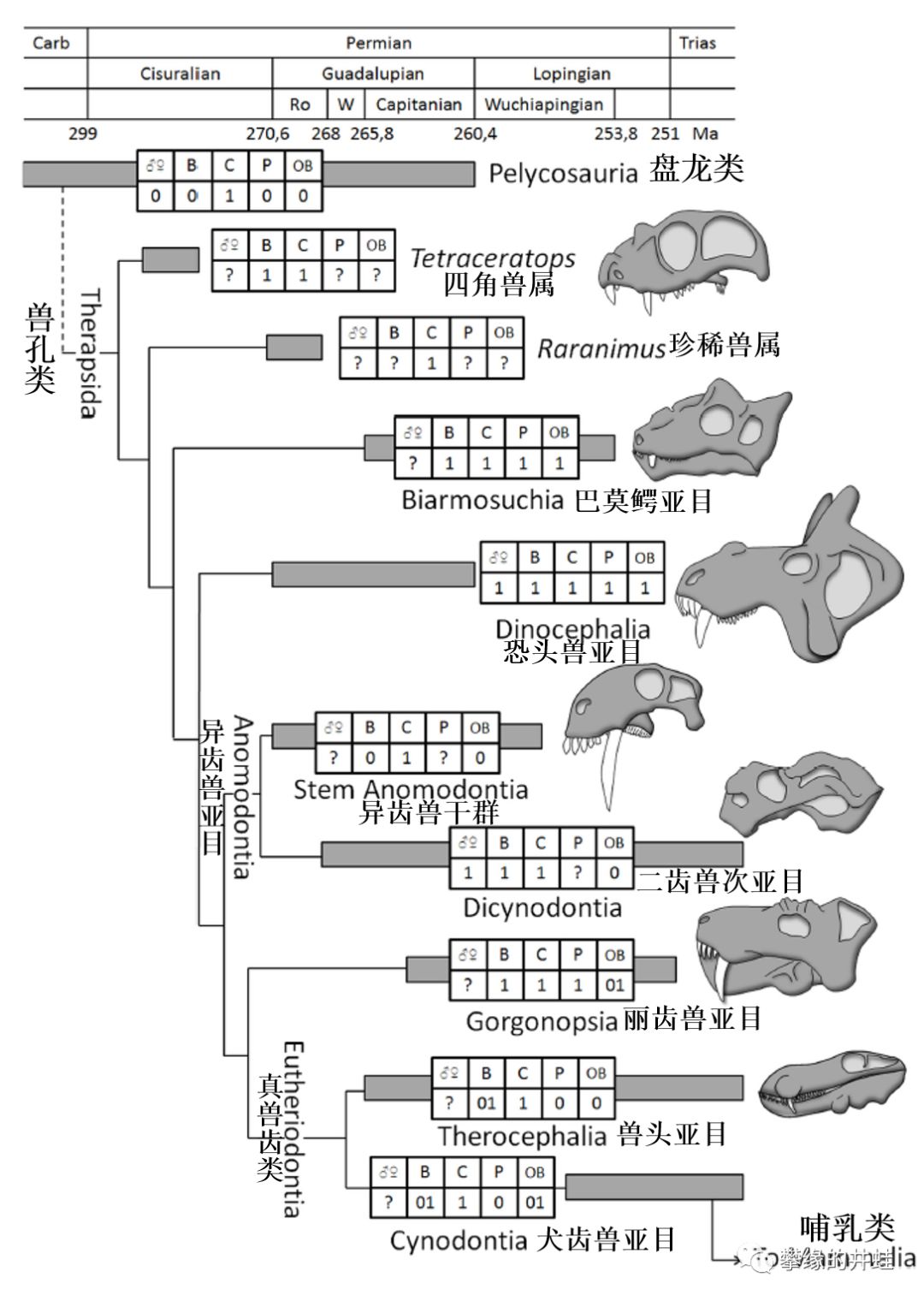
兽孔目主要类群和生存时代。四角兽和珍稀兽大致处在最基干的演化位置。大体上,越靠下的类群,演化程度越高。犬齿兽亚目包含一个演化支:哺乳形类Mammaliaformes,包括哺乳动物和最接近哺乳动物的类群。从图上也可以看出,发达的犬齿是大多数兽孔类的标配。图片来源自[6]。
牙齿分化和异型齿(Heterodont)的出现,极大提高了合弓纲的适应能力。要充分发挥异型齿的威力,需要有与之匹配的下颌结构和咬合肌牵引。关于兽孔目下颌的演化过程比较复杂,也比较漫长,本回只能起个头。
我们先来看一看早期羊膜动物的初始头骨构型:
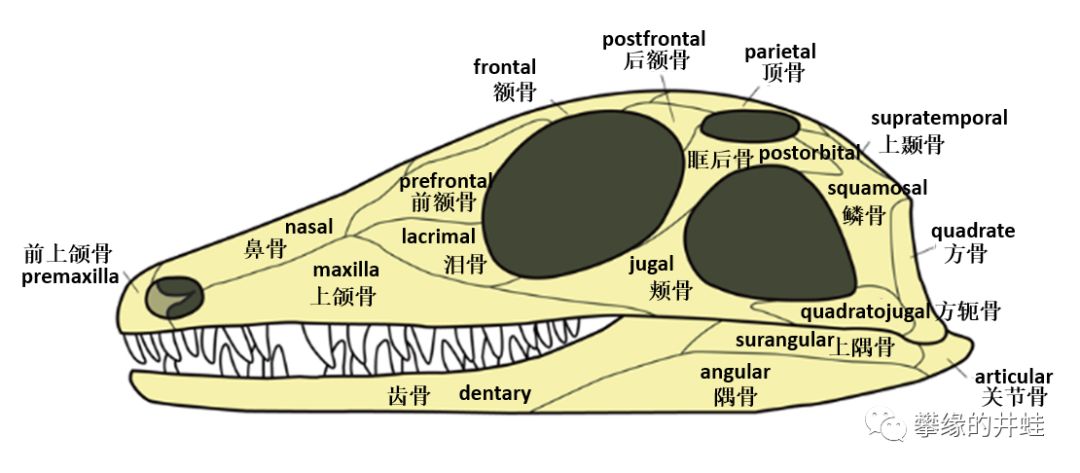
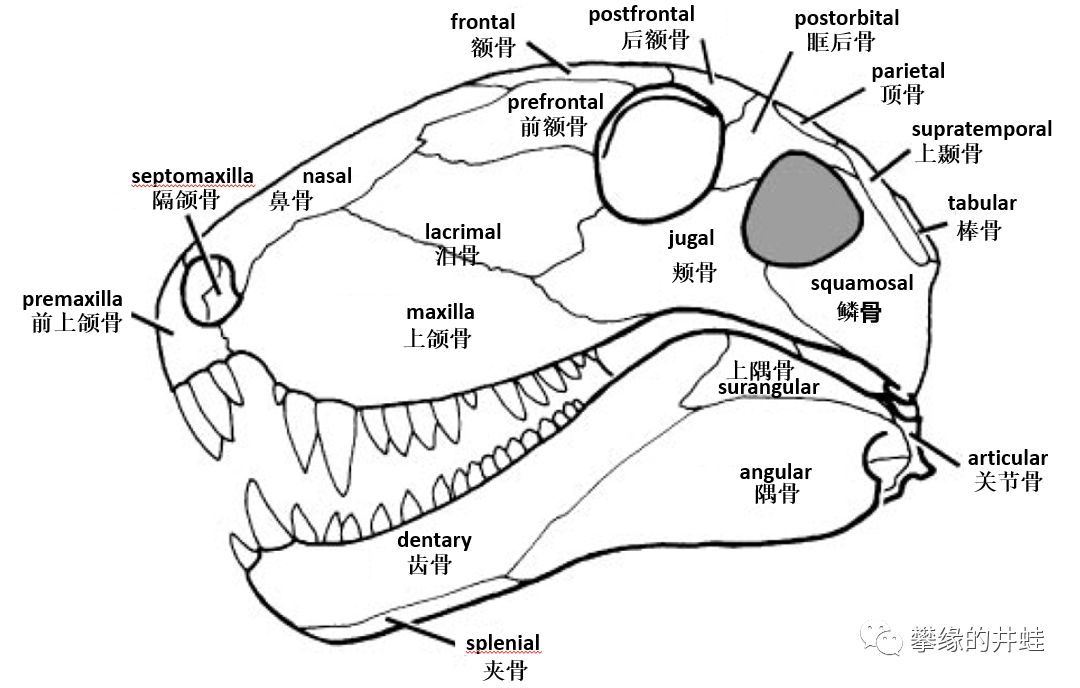
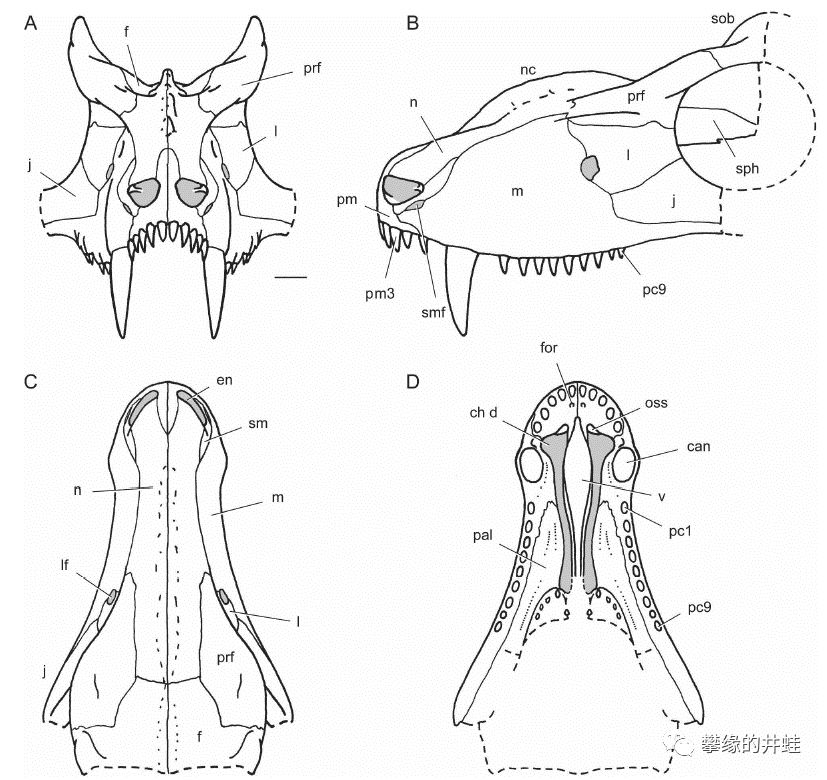
上图:杨氏蜥Youngina capensis(基干双孔类)的头骨结构(图片来源自[7]);下图:异齿龙的头骨结构(图片来源自[8])。除了颞颥孔的数量有区别,组成颅脑,前额和下颌的骨骼基本上一样。请注意组成下颌的几块骨头,从前到后依次是:齿骨,隅骨,上隅骨,和关节骨。
后来的蜥形纲基本上保持了这样的下颌结构,少有改变。

注意图中的下颌,齿骨,隅骨,上隅骨的分界非常明显。这种“拼接式”的下颌骨,还有形状功能基本一样的同型齿(Homodont),是蜥形纲头骨的主要特征。图片来源自网络。
而合弓纲对下颌进行了一系列魔改。主要是增强齿骨,退化其他骨骼。到最后隅骨和关节骨进到了耳朵里,上隅骨愈合进齿骨,下颌变成只由一块单独的齿骨构成。
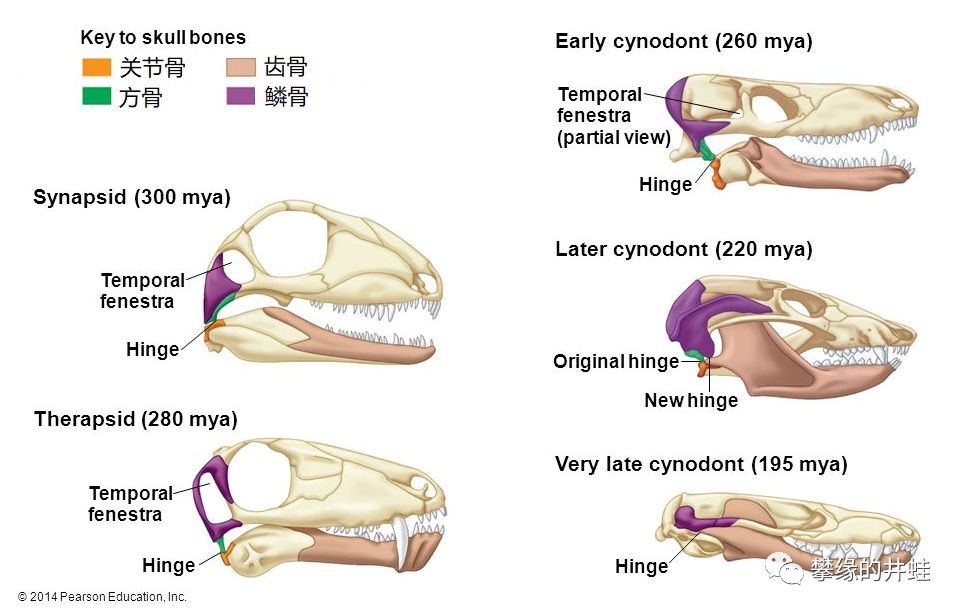
合弓纲下颌演化过程简图。可以看到兽孔目的演化阶段,齿骨不断强化,挤掉了其他下颌骨骼。图片来源见水印,有改动。

当合弓纲演化到哺乳动物的阶段,下颌只剩下一块齿骨,齿骨的末端嵌入上颌鳞骨形成的颞弓,形成特殊的齿骨-鳞骨关节(Dentary-SquamosalJoint)。图片来源自网络。


上图:沧龙头骨化石;下图:古蜥鲸头骨化石。尽管对趋同演化让两代海洋霸主呈现相近的外形,但根据蜥形纲的典型特征:同形齿和“组合式”下颌;合弓纲的典型特征:异型齿,单一下颌骨和齿骨-鳞骨关节,还是很轻易确定它们的归属。图片来源自网络。
和楔齿龙相比,兽孔目拥有更大的颞颥孔,可以容纳更加粗壮的肌肉。复合多种功能的异型齿,灵活的颌关节,配合强大的咬合力,让这些合弓兽的大嘴成为二叠纪最高效的进食工具,源源不断地收割泛古大陆的丰饶物产。
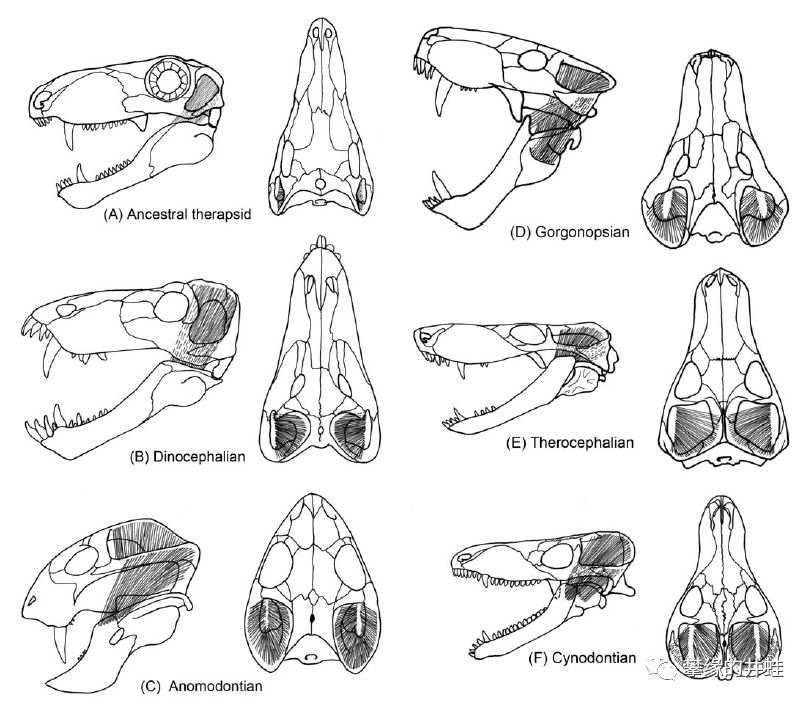
兽孔目主要类群的咬合肌结构复原图。A:巴莫鳄亚目;B:恐头兽亚目;C:异齿兽亚目;D:丽齿兽亚目;E:兽头亚目;F:犬齿兽亚目。图片来源自[3]。
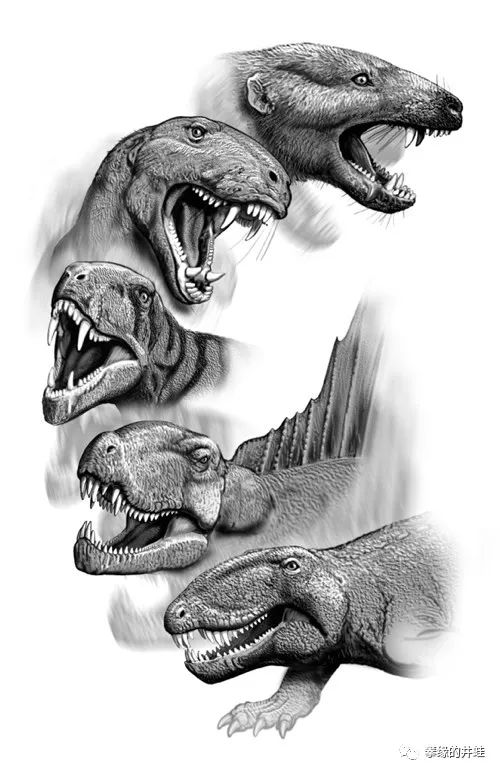
盘龙目到兽孔目到兽孔目头部演变示意图。合弓纲在二叠纪真正实现了改头换面,从“龙头”变成“兽头”。图片来源自网络。
除了四角兽和珍稀兽之外,比较接近兽孔目基干类群的还有巴莫鳄亚目(Biarmosuchia)。这是一个很庞杂的并系群,大多是中型肉身性动物,少数种类大型。
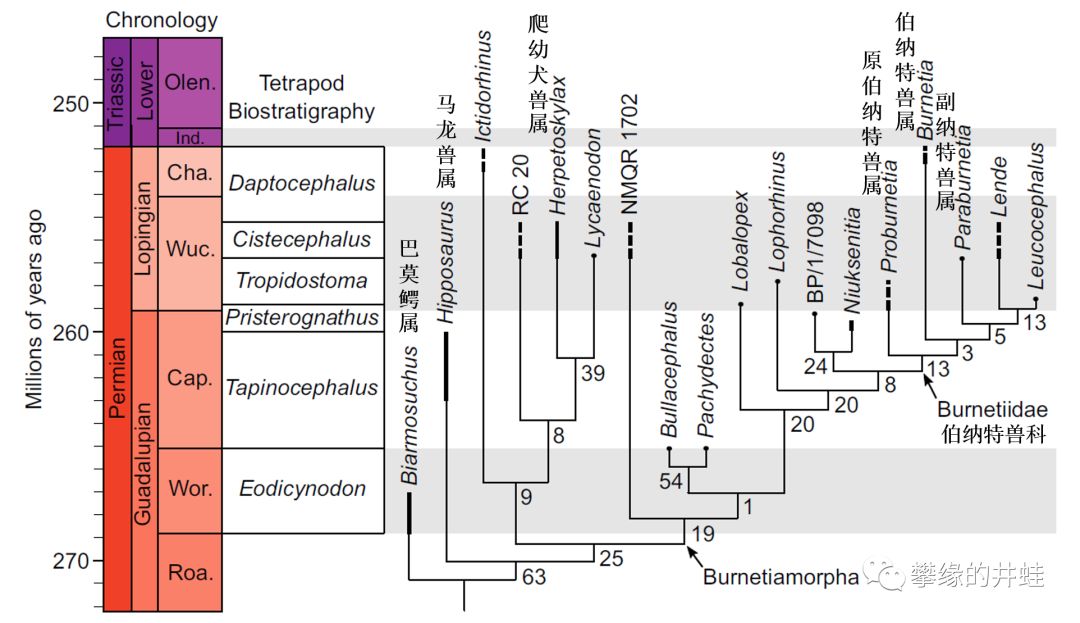
巴莫鳄亚目的演化谱系。早期种类以巴莫鳄属为代表,后期出现了伯纳特兽类这样特化的类群。图片来源自[9]。
虽然名字里有个“鳄”字,但无论外形还是生活方式,巴莫鳄都一点也不像鳄鱼,亲缘关系更是差了十万八千里。从仅有的躯体和四肢化石来看,巴莫鳄类拥有苗条的身形和修长的四肢,适合快速奔行。
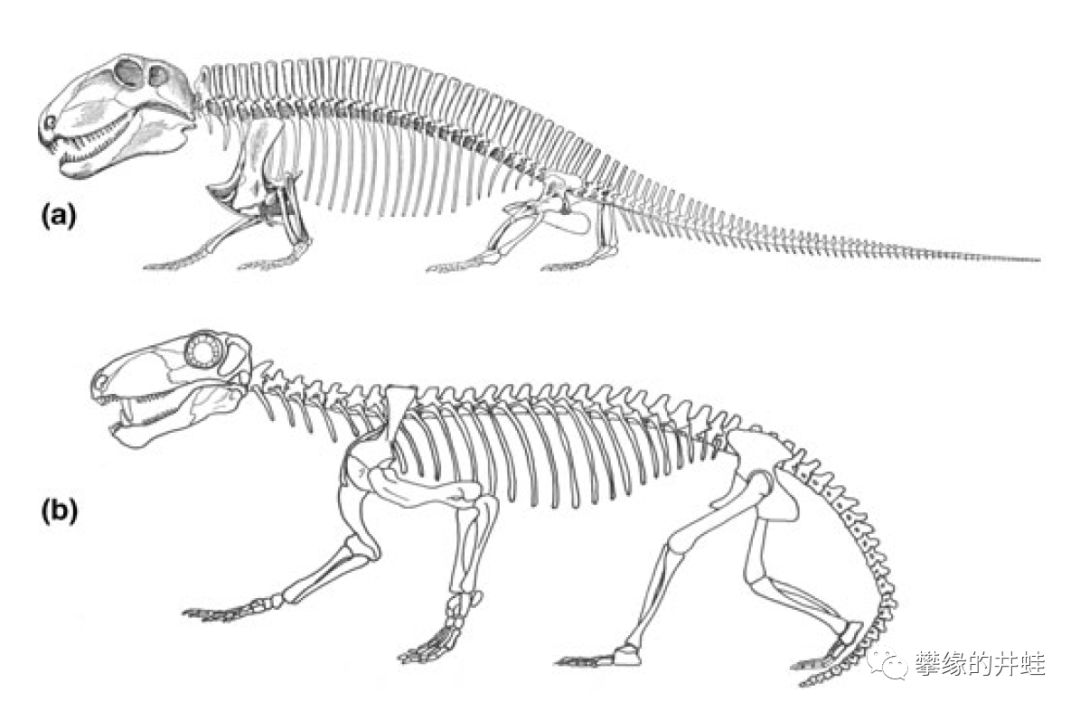
楔齿龙(a)和巴莫鳄(b)骨骼复原图。巴莫鳄的头骨结构仍然和楔齿龙近似,颞颥孔,异型齿和下颌齿骨都有进一步的演化。肢骨更长而粗壮,可以把身体抬离地面。看看两者的对比,就不难理解为什么盘龙目会退出历史舞台了。遗憾的是,由于早期化石资料的缺失,我们还无从知道兽孔目是怎样从“蜥蜴爬”过渡到“直立跑”。图片来源自[2]。

俄罗斯彼尔姆地区(Perm region)发现了丰富的早期巴莫鳄化石,比如上图的Biarmosuchus tener。它可能是人们了解得最深入的巴莫鳄了。图片来源自网络。
成体Biarmosuchus tener全长估计在2.5米左右,曾经被命名为另一种动物:始巨鳄(Eotitanosuchus olsoni)。听名字很容易让人想起电影里的食人怪物,但实际上并没有那么恐怖。
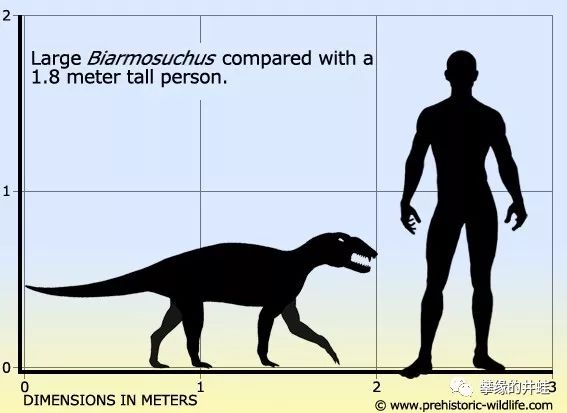
2米长的大型巴莫鳄。这么看起来,2.5米的“始巨鳄”大概也就是一条阿拉斯加的大小。整个巴莫鳄亚目就没有演化出顶级掠食者。图片来源见水印。
南非的卡鲁盆地(Karoo Basin)和附近地区发现了许多演化后期的巴莫鳄类化石,但是基本上都只留下了头骨(当然很可能是井蛙没有找到,如果有大神有这些巴莫鳄的骨架照片,可否分享一下,不胜感谢),比如这些:
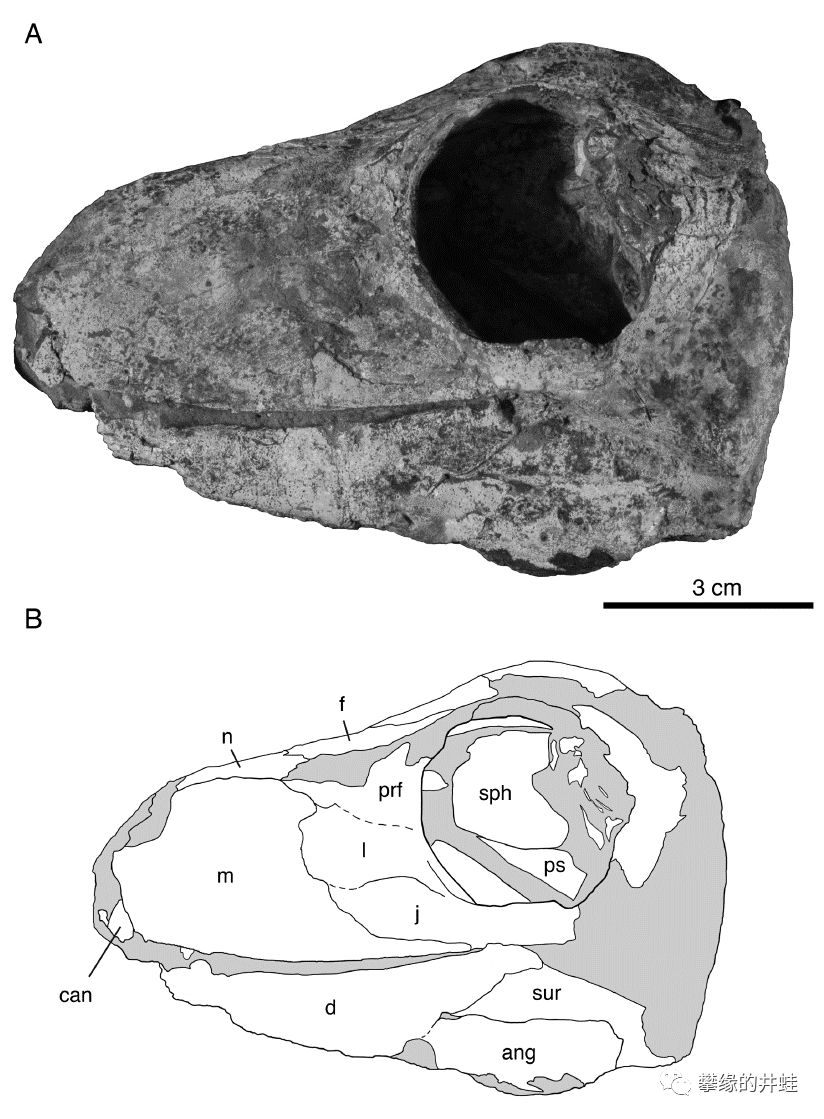
赞比亚琅瓜盆地(Luangwa Basin)发现的巴莫鳄类头骨化石。这里发现的二叠纪动物群和卡鲁盆地的非常接近。图片来源自[10]。
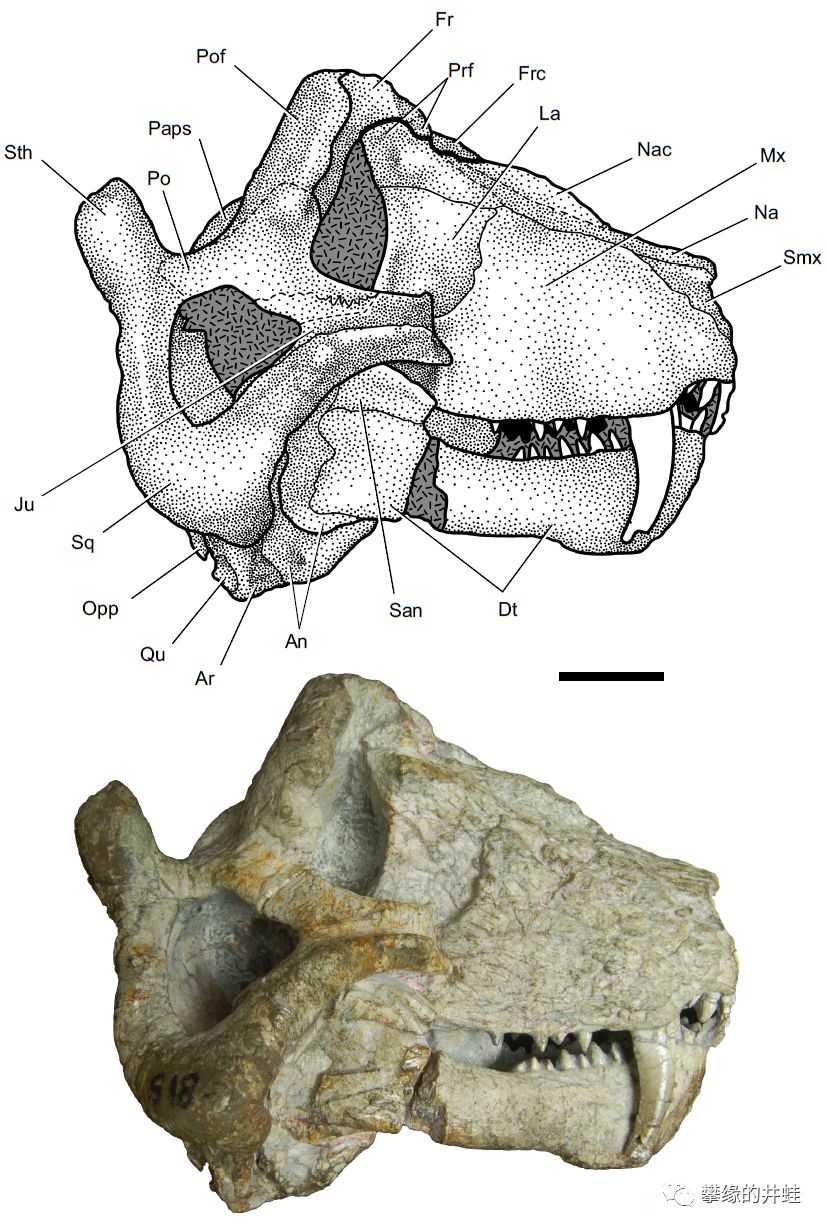
卡鲁盆地发现的巴莫鳄类Leucocephalus wewersi化石。图片来源自[9],标尺长度2厘米。
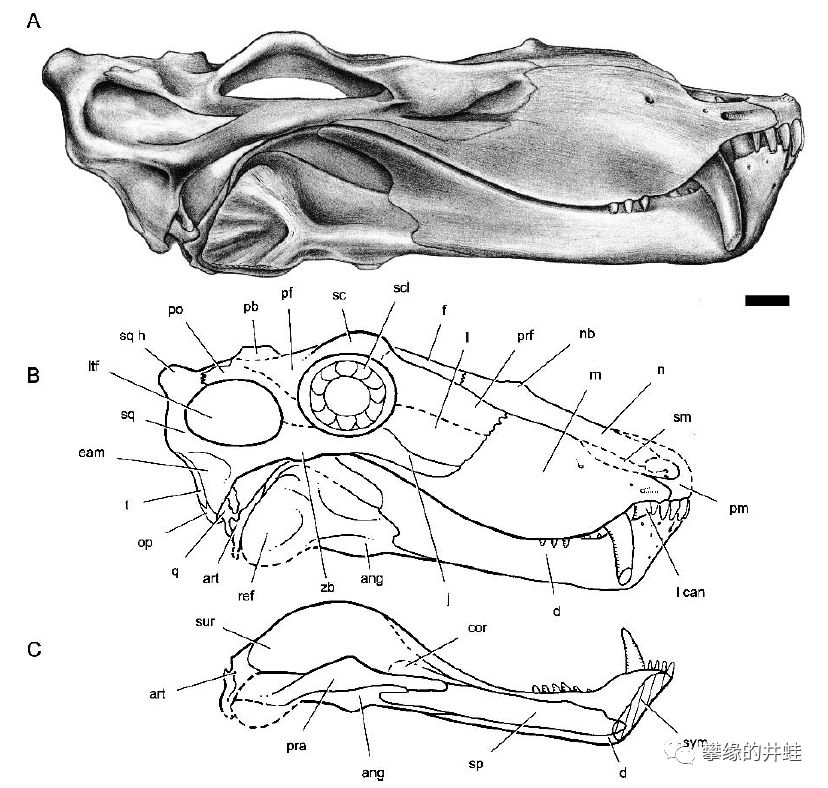
卡鲁盆地发现的巴莫鳄类Lobalopex mordax化石。图片来源自[11],标尺长度1厘米。

卡鲁盆地还发现了一只巴莫鳄Baby的完整头骨。它们小时候居然是个大眼萌,标志性的犬齿也还没有长出来,还蛮可爱的。图片来源自[12]。

研究这认为这个Baby可能是Lemurosauruspricei(右)的幼体。岁月果然是把杀猪刀啊。图片来源自[12]。
上面说到的Leucocephalus,Lobalopex,Lemurosaurus都属于“伯纳特兽形类Burnetiamorpha”。是巴莫鳄演化后期的一个比较重要的分支。在它们身上(其实也就是头上),已经出现了一些装饰,比如突出的眉弓,还有头骨上的瘤结。
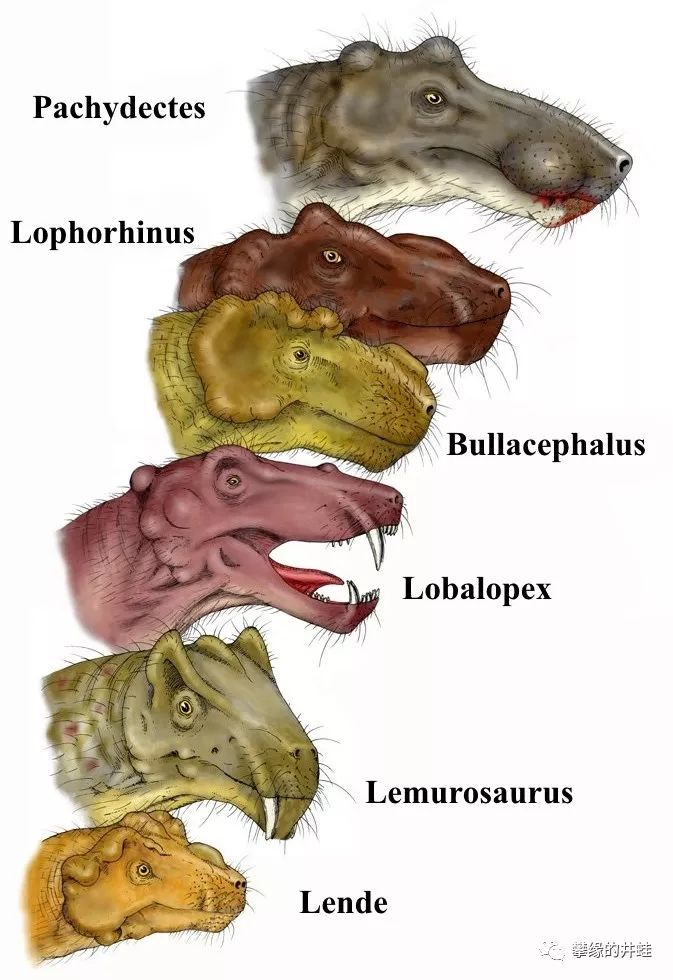
一些伯纳特兽类的头部复原图。它们的脑袋变得疙疙瘩瘩的。看不出这些结构对狩猎或者争斗有什么帮助,很可能是用来向异性展示魅力的第二性征。图片来源自网络。
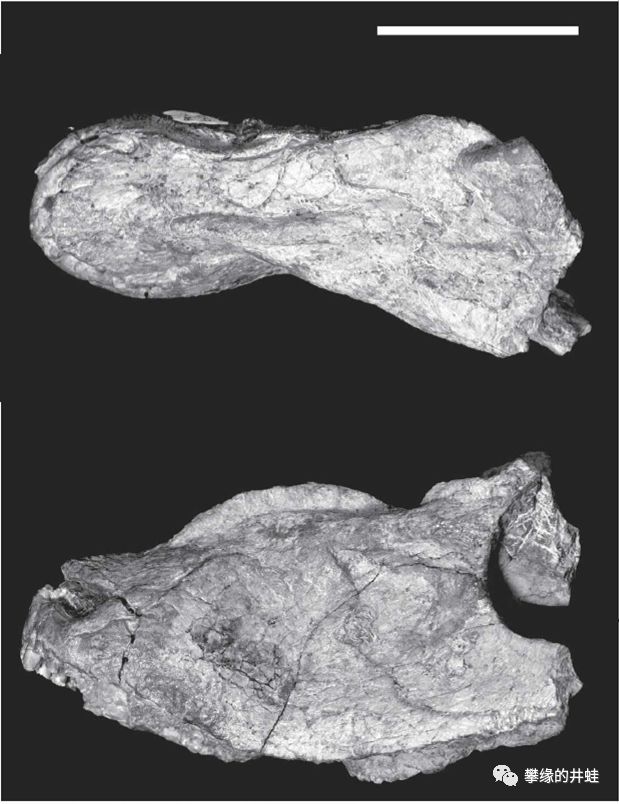
卡鲁盆地发现的另一种伯纳特兽类:Lophorhinus willodenensis。它的眉弓特别突出,鼻骨正中还有一道隆起的中脊。图片来源自[13],标尺长度4厘米。

Lophorhinus willodenensis正脸复原图。看起来甚是凶悍。图片来源自网络。
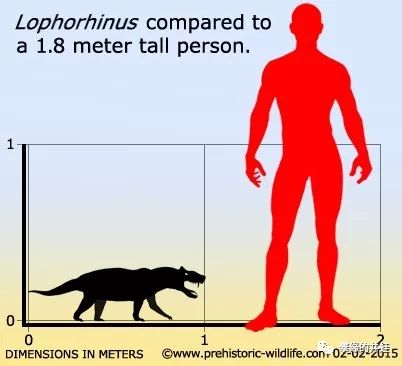
Lophorhinus willodenensis的实际大小。伯纳特兽类基本上都是这个体型。但是别把这个小东西看扁了:锋利的犬齿,强大的咬合力,灵活的身体加上直立奔跑的速度优势,让它足以猎杀比自己大得多的动物。图片来源见水印。
总体来说,巴莫鳄亚目长得都比较“中规中矩”,无论体型还是特化的装饰结构,都远远没有其他兽孔目类群那么夸张。很可能是由于身体结构相对原始,巴莫鳄类一直都不算繁盛。身为食肉兽的它们从来都没能攀上陆地食物链的顶峰。“始巨鳄”昙花一现,很快就被恐头兽亚目和丽齿兽亚目的超级掠食者驱逐。二叠纪的泛古大陆迎来了自己的虎豹熊罴。异齿兽亚目和另一些恐头兽类则演化成了二叠纪的犀象牛马。生存到二叠纪晚期的伯纳特兽类都是一些中小型种类,大约只能扮演着类似豺狼獾鼬一类的角色。然而在二叠纪结束前的500万年间,另一个合弓纲类群:犬齿兽亚目迅速崛起,这些小型食肉兽比伯纳特兽类更凶悍,更敏捷,演化程度更高,适应能力更强。在犬齿兽亚目的冲击下,巴莫鳄亚目很快凋零。之后,惨烈的大灭绝事件埋葬了最后的孑遗。
地球名片
生物分类:动物界-脊索动物门-脊椎动物亚门-羊膜类-合弓纲-兽孔目-巴莫鳄亚目
存在时间:二叠纪中期至二叠纪晚期
现存种类:无
化石种类:未找到完整统计
生活环境:陆地
代表特征:中到大型食肉动物,头骨结构接近楔齿龙,颞颥孔更大,牙齿分化更明显
包含种类:巴莫鳄,马龙兽,爬幼犬兽,伯纳特兽,原伯纳特兽,副伯纳特兽
参考文献
[1] Neil Brocklehurst, Christian F. Kammerer, Jörg Fröbisch,The early evolution of synapsids, and the influence of sampling on their fossilrecord. The Paleontological Society, Paleobiology, 39(3): 470-490, 2013, DOI:http://dx.doi.org/10.1666/12049
[2] T. S. KEMP, The origin and early radiation of thetherapsid mammal-like reptiles: a palaeobiological hypothesis. THE AUTHOR 19(2006), 1231–1247, doi: 10.1111/j.1420-9101.2005.01076.x
[3] Tom S. Kemp, Phylogenetic interrelationships and patternof evolution of the therapsids: testing for polytomy. Palaeont. afr. (December2009), 44: 1–12
[4] Amson, E., Laurin, M., On the affinities ofTetraceratops insignis, an Early Permian synapsid. Acta PalaeontologicaPolonica 56 (2): 301–312, 2011
[5] Liu, J., Rubidge, B., and Li, J., New basal synapsidsupports Laurasian origin for therapsids. Acta Palaeontologica Polonica, 54(3): 393–400, 2009, DOI: 10.4202/app.2008.0071.
[6] Benoit J, Manger PR, Fernandez V, et al., Cranial Bossesof Choerosaurus dejageri (Therapsida, Therocephalia): Earliest Evidence ofCranial Display Structures in Eutheriodonts. PLoS ONE 11(8): e0161457, 2016,doi: 10.1371/journal.pone.0161457
[7] Robert L. Carroll, Plesiosaur Ancestors from the UpperPermian of Madagascar. Philosophical Transactions of the Royal Society ofLondon, Series B (Biological Sciences). 293: 315-383, 1981, doi:10.1098/rstb.1981.0079
[8] Bruce S. Rubidge,Christian A. Sidor, Evolutionary Patterns among Permo-Triassic Therapsids.Annual Review of Ecology and Systematics, Vol. 32 (2001), pp. 449-480
[9] MICHAEL O. DAY, ROGER M. H. SMITH, JULIEN BENOIT, etal., A NEW SPECIES OF BURNETIID (THERAPSIDA, BURNETIAMORPHA) FROM THE EARLY WUCHIAPINGIANOF SOUTH AFRICA AND IMPLICATIONS FOR THE EVOLUTIONARY ECOLOGY OF THE FAMILYBURNETIIDAE. Papers in Palaeontology, 2018, pp. 1–23, doi: 10.1002/spp2.1114
[10] Christian A. Sidor, The first biarmosuchian from theupper Madumabisa Mudstone Formation (Luangwa Basin) of Zambia. PalaeontologiaAfricana, 49: 1–7
[11] CHRISTIAN A. SIDOR, JAMES A. HOPSON, ANDRÉ W. KEYSER, ANEW BURNETIAMORPH THERAPSID FROM THE TEEKLOOF FORMATION, PERMIAN, OF SOUTHAFRICA. Journal of Vertebrate Paleontology, 24(4): 938–950, 2004
[12] Aliénor Duhamel, Julien Benoit, Juvenile Biarmosuchiansfrom the Karoo Beaufort Group Shed New Light on Basal Therapsid Ontogeny.Presentation, July 2018, DOI: 10.13140/RG.2.2.36435.48166
[13] CHRISTIAN A. SIDOR, ROGER M. H. SMITH, SECONDBURNETIAMORPH THERAPSID FROM THE PERMIAN TEEKLOOF FORMATION OF SOUTH AFRICA ANDITS ASSOCIATED FAUNA. Journal of Vertebrate Paleontology, 27 (2):420–430, 2007