

这一回,井蛙要试着梳理一下自己的族谱了。
两栖动物(Amphibian)因为分布广泛,数量众多,味道鲜美,营养丰富,与农业生产关系密切而为人们熟知。从“稻花香里说丰年,听取蛙声一片”,到“荷花上面有蛤蟆,一戳一蹦跶”,到启蒙动画《小蝌蚪找妈妈》,再到满世界的牛蛙馆子,都是人类和两栖动物亲密关系的证明。
所有现代两栖动物都属于滑体两栖类(Lissamphibia),顾名思义,它们的皮肤裸露,皮下布满腺体,分泌大量粘液,所以摸上去总是冰冷湿滑的。滑体两栖动物分为三个目:无尾目(Anura),包括各种蛙和蟾蜍;有尾目(Caudata),包括各种鲵和蝾螈;无足目(Gymnophiona),包括各种鱼螈和蚓螈。每个目的特征也都直白地写进了名字里。

相比无尾目和有尾目,无足目尤其稀少而神秘。我国只有两种无足目两栖动物:双带鱼螈Ichthyophis glutinosus和版纳鱼螈Ichthyophis bannanicus。这两种鱼螈的外形非常相似,井蛙也不知道上面照片里的是哪种。图片来源见水印。
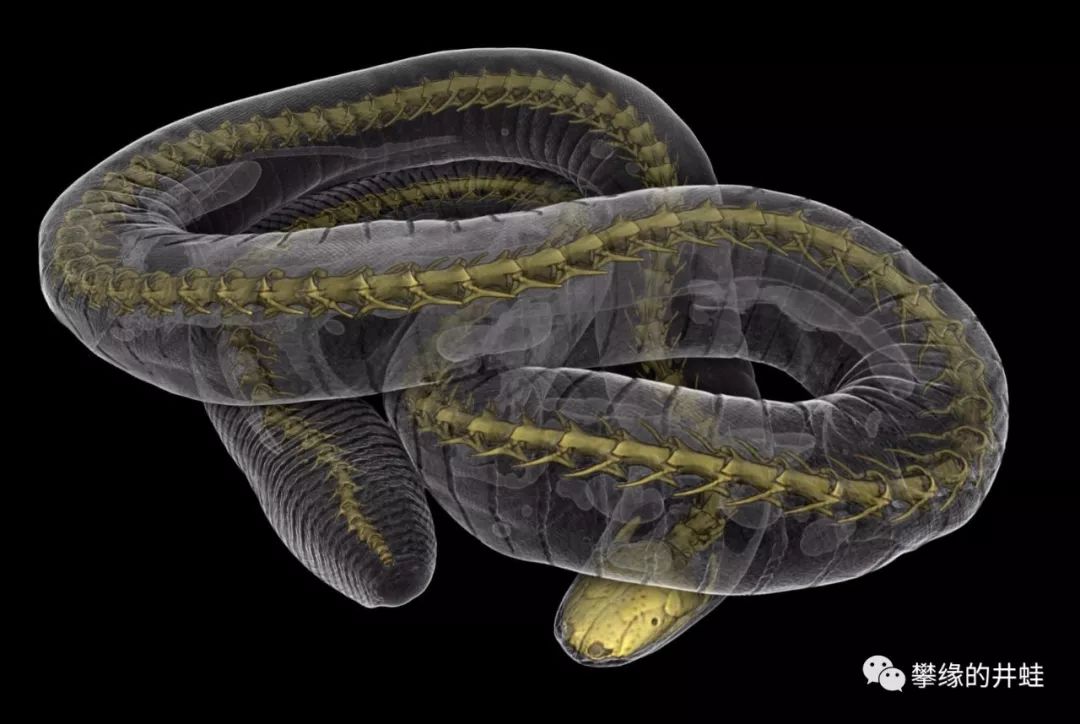
无足目四肢完全消失,眼睛退化,柔软湿滑的皮肤上环布一节一节的皱褶,样子非常像一条大蚯蚓。但它的身体里是有一根发育完全的脊椎骨的。图片来源自网络。
滑体两栖类主要占据水陆之间的一些偏僻的生态位,与此相应地,它们身体结构也都极端特化:无尾目退化了尾巴,无足目退化了眼睛和四肢,一些有尾目甚至不再长出肺。
所有的这一切,和地层中发现的那些庞大,笨重,凶猛,怪异的“古两栖动物(主要指离片椎类和壳椎类)”化石相比,是那么的格格不入。
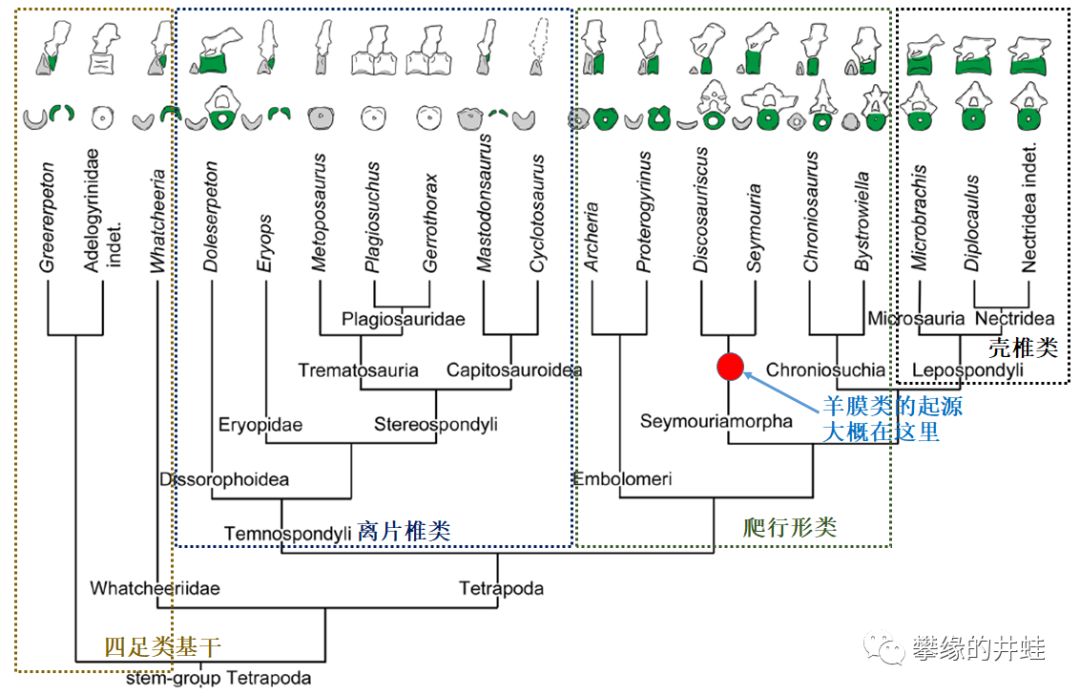
“古两栖动物”的演化谱系和它们的脊椎结构。井蛙标出了羊膜类大概的位置,所以从系统发生学上看,壳椎类其实更接近羊膜动物。图片来源自[1]。


大体上,“古两栖动物”包括四足类基干(Stem Tetrapods),离片椎类(Temnospondyli)和壳椎类(Lepospondyli)。上图是引螈Eryops(离片椎类)的骨骼。下图是笠头螈Diplocaulus(壳椎类)的骨骼。它们都有发育完全的脊柱,强大的肋骨和肢骨,尤其是头骨非常坚固厚实,看上去充满力量。图片来源自网络。
然而滑体两栖类的骨骼却是这样的:

无尾目成体的尾骨消失,脊柱数量减少,前肢桡骨与尺骨愈合成桡尺骨,后肢胫骨与腓骨愈合成胫腓骨。这些改造让蛙类短小精悍,既能上岸跳跃,也能入水蛙泳。图片来源自网络。

有尾目和无尾目一样,前肢只有四个脚趾。脊柱和肢骨纤细薄弱,肋骨退化,不足以支撑胸腔,所以这些动物都是软趴趴的。图片来源自网络。

足目的骨骼,四肢退化得一点痕迹都不剩。蛇鳝一般缠长的身体适合在泥土和洞穴中穿行,捕食蠕虫和甲壳动物。图片来源自网络。
除了无足目的头骨还算完整外,无尾目和有尾目的头骨都高度简化,大量骨片愈合或者消失,两个眼洞的比例大得出奇。所以它们的头骨显得很“碎”很“空”,完全不同于古两栖动物那种一整块大厚板的结构。
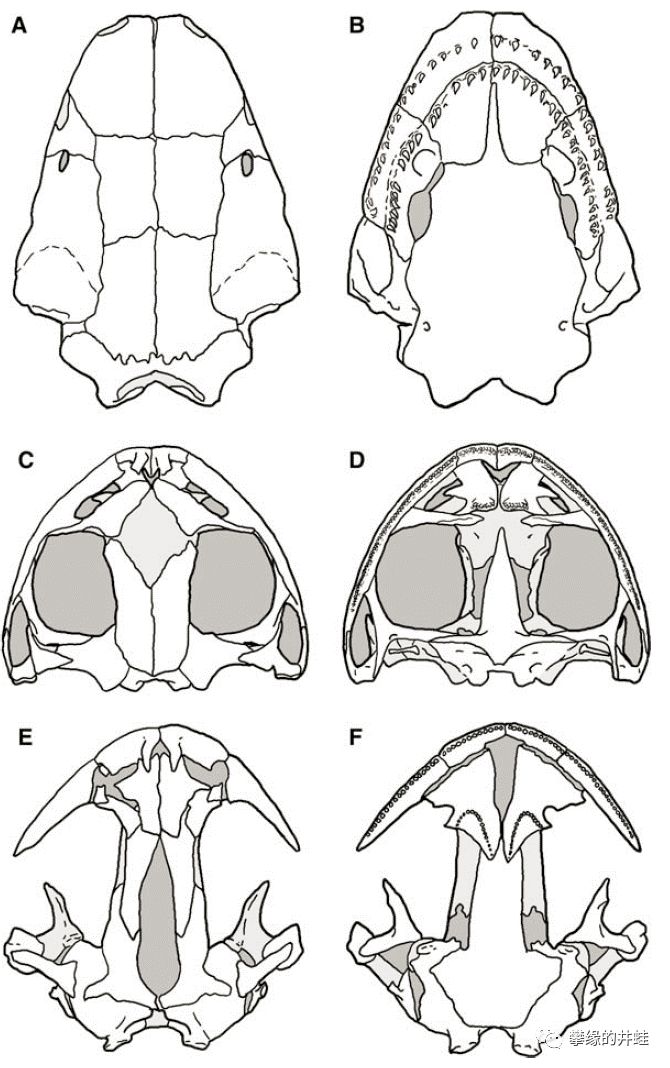
滑体两栖类的颅骨结构。A,B:墨西哥蚓螈Dermophis mexicanus(无足目)。C,D:南美囊蛙Gastrotheca walkeri(无尾目)。E,F:极北鲵Salamandrella keyserlingii(有尾目)。图片来源自[2]。
和古两栖类一比,滑体两栖类简直就是一群软骨头的不肖子孙。但问题是,井蛙到底应该去压住谁的棺材板呢?离片椎类,还是壳椎类的呢?
古生物研究者一直对滑体两栖类的起源争论不休。因为脊椎结构接近(参见第一百九十一回Fish to Beast),所以有人坚持离片椎类是滑体两栖类的祖宗——离片椎类起源说。也有人说滑体两栖类的外形如此多样,一定是壳椎类的后裔(参见第一百九十二回Normal to Bizarre)——壳椎类起源说。还有人认为无尾目和有尾目起源自离片椎类,而无足目起源自壳椎类——多源说。嘿,能说的话全让你们说了。
幸好我们还有分子生物学这个最有力的武器。根据对现存两栖动物的基因测序对比,得到了以下结论:
1.滑体两栖类是一个单系群。并没有出现“明明是个蛙却和鳄鱼亲缘关系更近”这种尴尬的局面。
2.无尾目,有尾目和无足目各自也都是单系群。并没有出现“明明是个蛙却和娃娃鱼亲缘关系更近”这种尴尬的局面。
3.无尾目和有尾目的亲缘关系更近,它们组成了一个演化支(蛙形总目Batrachia)。而无足目是一个早早分化出去的旁支。
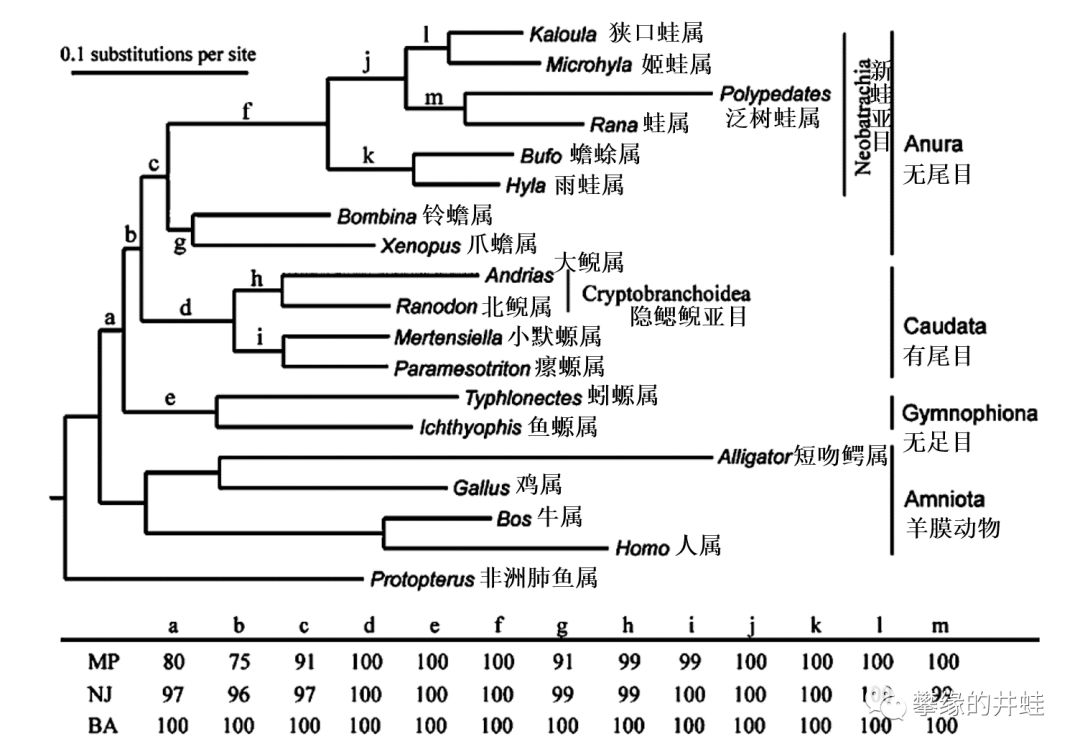
根据线粒体基因序列(Mitochondrial genome sequences)分析绘制的滑体两栖类谱系图。所有的滑体两栖类都可以追溯到一个共同祖先(a),然后无足目先分化出去(e);之后某个共同祖先(b)的后代分化出无尾目和有尾目两支。图片来源自[3]。
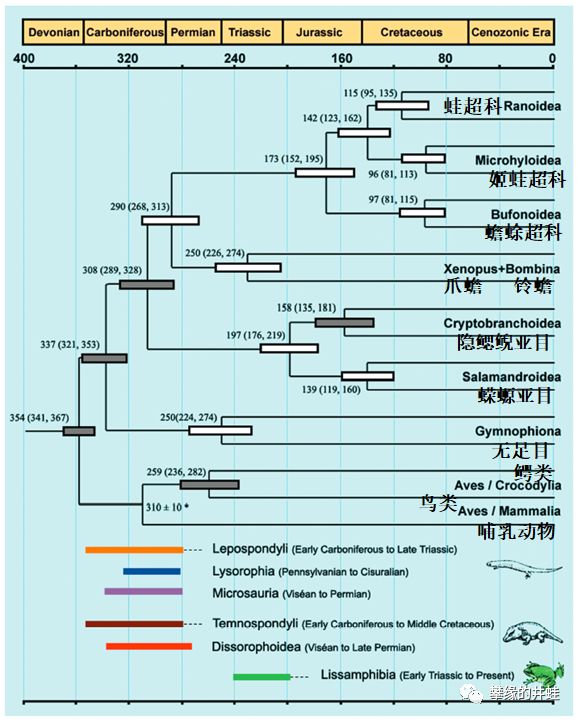
根据分子钟推出的现存四足动物谱系和分化时间。根据这项研究,滑体两栖类最近的共同祖先大概生活在石炭纪早期。图片来源自[3]。
这样一来,基本可以排除壳椎类起源说和多源说。只剩下离片椎类起源说,但还要接受进一步的验证。
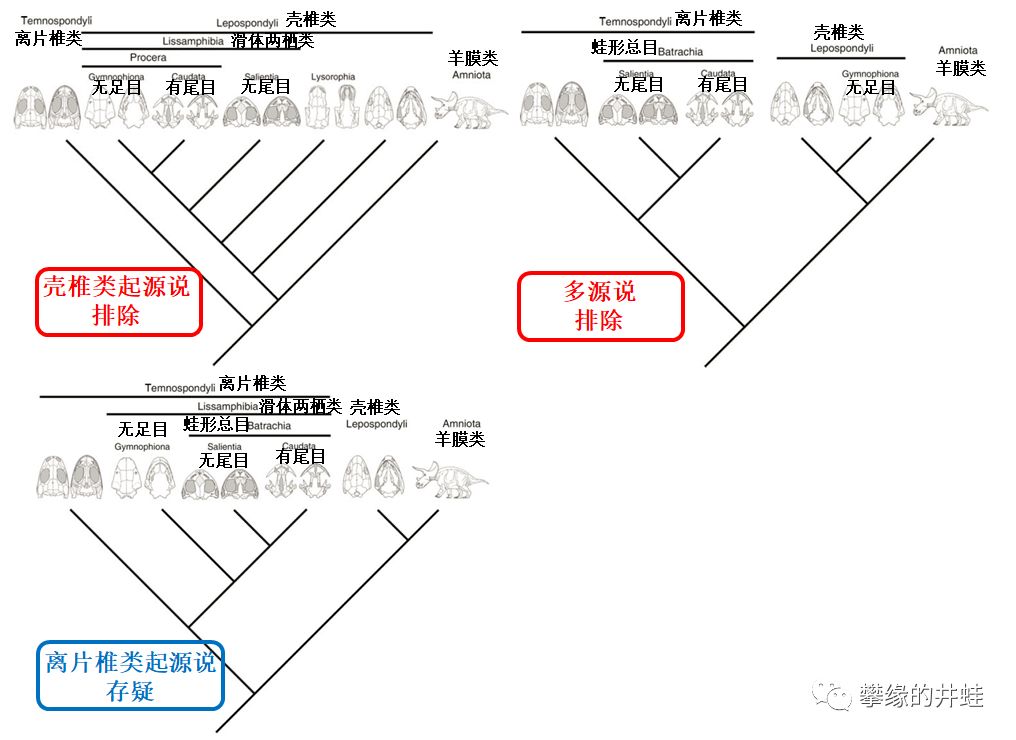
关于滑体两栖类起源的三种假说。分子生物学方法排除了两种。如果离片椎类起源说真的成立,那到底哪一类离片椎类才是滑体两栖类的祖先呢?图片来源自[2]。
接下来轮到比较解剖学登场。滑体两栖动物会长出一种非常特殊的牙齿:Pedicellate teeth(可能翻译成茎齿?梗齿?管齿?请两栖类方面的专家指正)。
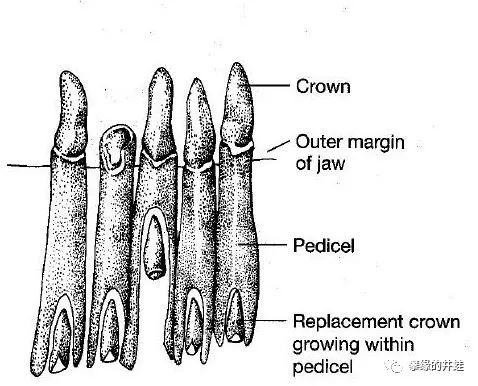
Pedicellate teeth的结构。齿冠下面的部分套在一根长长的管子(Pedicel)里,这根管子深埋进颌骨内部。新的齿冠就在管子里成长,上升,最后顶出管口,取代旧的齿冠。图片来源自网络。
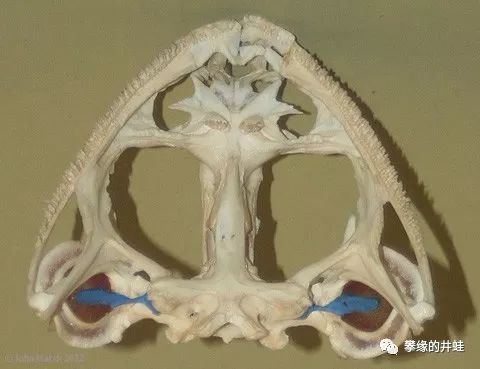

上图:美国牛蛙Rana catesbeiana的头骨;下图:美洲大鲵Cryptobranchus allegheniensis的头骨。注意颌骨上细密的小尖牙,这些都是Pedicellate teeth。图片来源自网络。
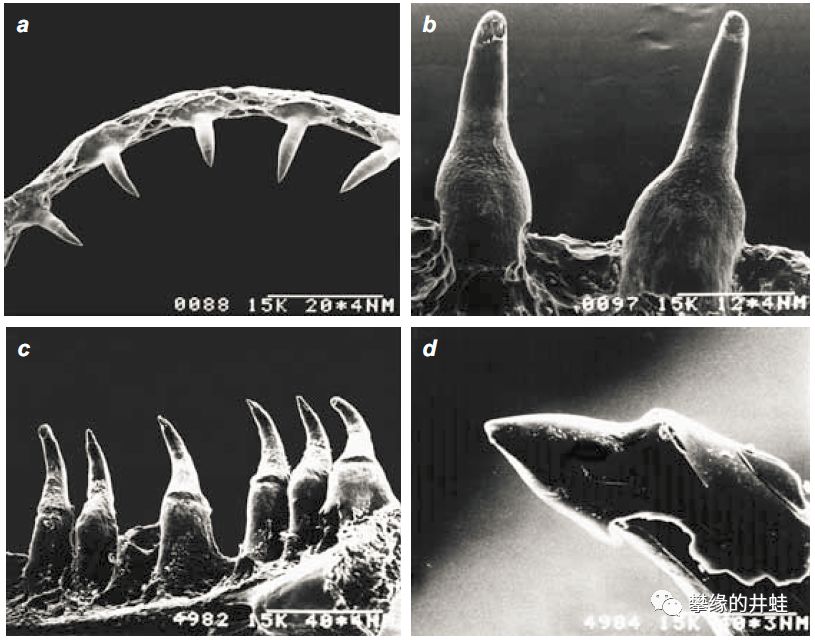
新疆北鲵Ranodon sibiricus幼体Pedicellate teeth的发育过程。早期幼体的牙齿没有套管子(a),长到亚成体之后,逐渐套上管子变成Pedicellate teeth(b到c)。图片来源自[4]。


新疆北鲵的样子。是不是非常可爱?图片来源见水印。
除了滑体两栖类,Pedicellate teeth只出现在一些小型离片椎类——双疏螈类Dissorophoidea的嘴里。这个证据,更加夯实了离片椎类起源说。
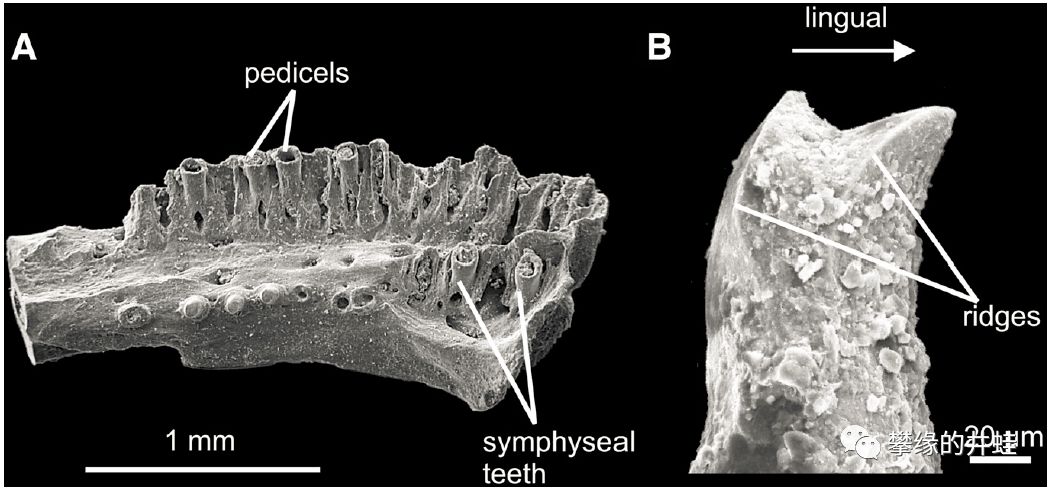
一种早二叠世双疏螈类Doleserpeton的Pedicellate teeth。A是嵌在颌骨上的管子;B是伸出管子的齿冠。图片来源自[5]。
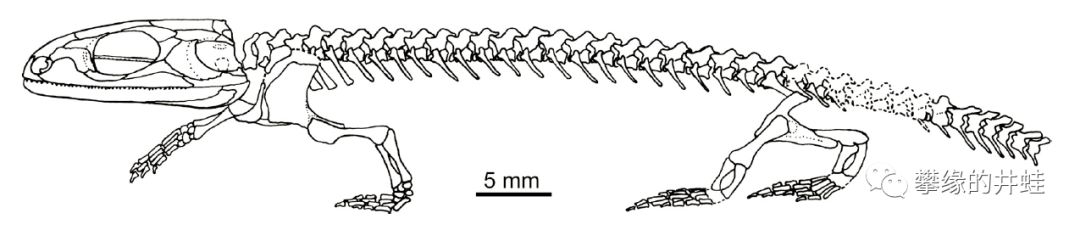
Doleserpeton骨骼复原图。它的外形像一条蝾螈,但是骨骼,尤其是头骨,比现代两栖类结实多了。图片来源自[5]。
随着越来越多的双疏螈类化石被发现,一些种类的头骨结构似乎也更加接近滑体两栖动物,向着更加轻薄的方向发展。
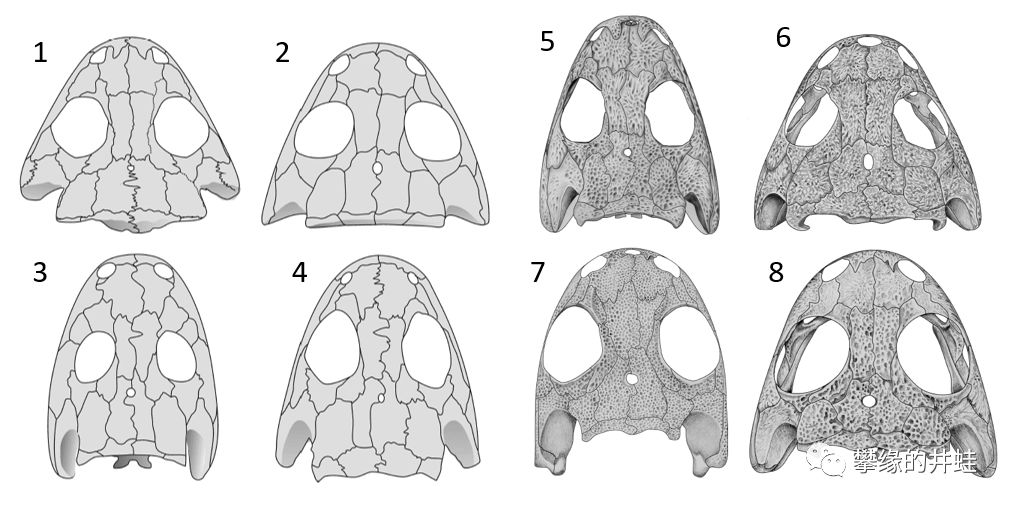
一些双疏螈类头骨结构。1:Limnogyrinus elegans,头骨长1.8厘米。2:Branchierpeton amblystomum,头骨长2厘米。3:Perryella olsoni,头骨长4.1厘米。4:Eimerisaurus graumanni,头骨长3.2厘米。5:小合螈Micromelerpeton credneri,头骨长4厘米。6:幻螈Apateon caducus,头骨长2.9厘米。7:两栖螈Amphibamus grandiceps,头骨长1.8厘米。8:幻螈Apateon gracilis,头骨长1.8厘米。这些小型离片椎类的眼睛越来越大,脑袋越来越浑圆,也越来越轻巧,逐渐接近蛙和蝾螈的样子。图片来源自[6]。
这些双疏螈类的外形已经很像现代有尾目了:圆头,大眼,扁身,四条小短腿,宽大的尾巴,可能还有终生保留的外鳃。我们甚至还能知道它们身上的花纹是什么样子。
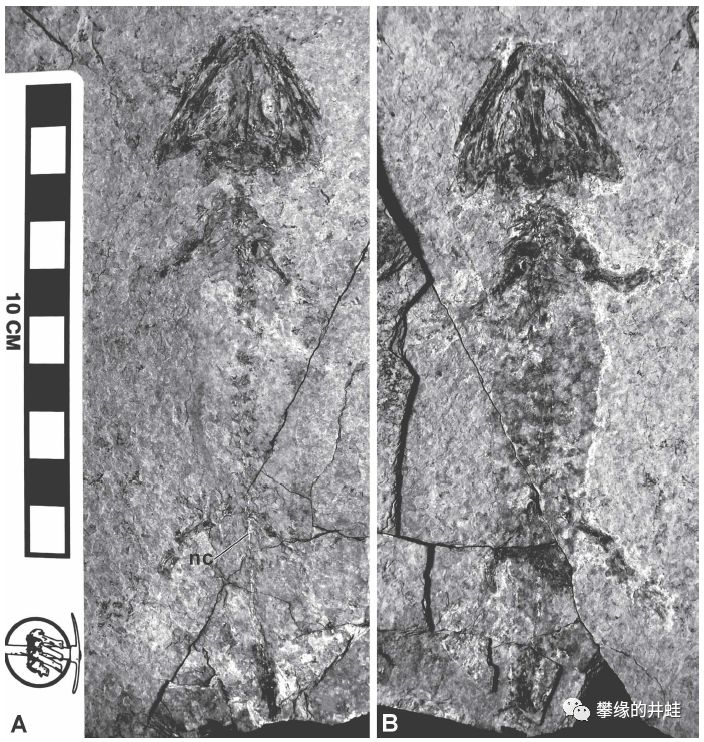
德国萨克森盆地(Saxony Basin)发现的早二叠世双疏螈类Melanerpeton tenerum化石。这个珍贵的标本保留了皮肤上的色素分布,可以看到腹侧(A)没有花纹,而背侧(B)有圈圈点点的花斑。图片来源自[7]。
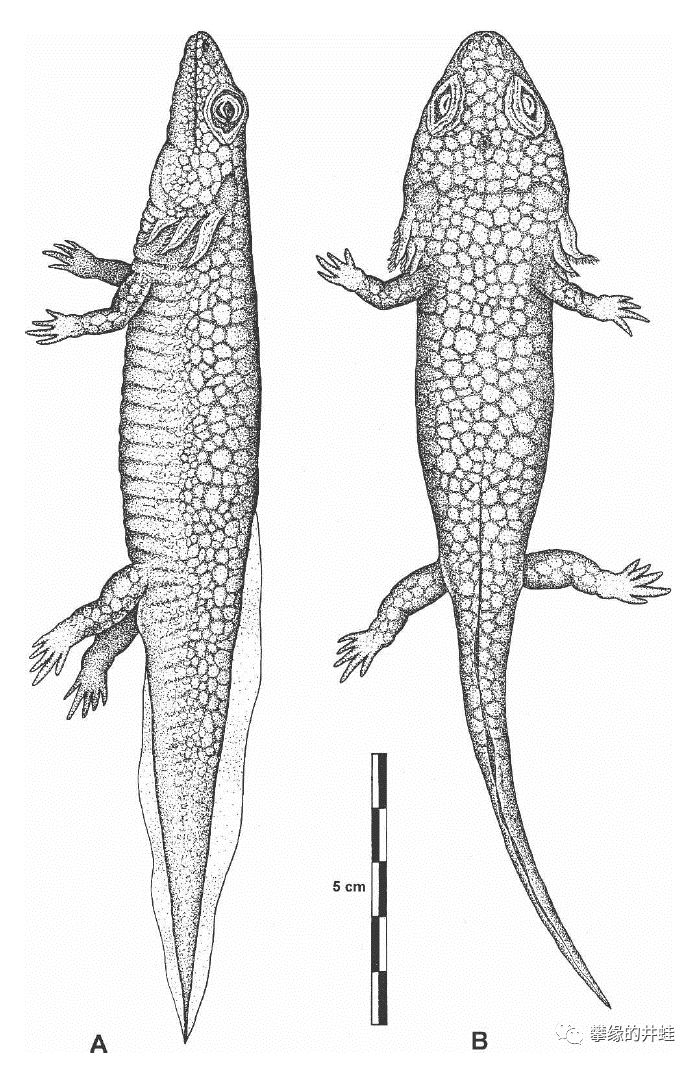
Melanerpetontenerum复原图。它的背部有大理石一样的花纹,也许有助于隐蔽在河水的波光中。图片来源自[7]。
但是双疏螈类和滑体两栖类还有一个巨大的差别:它们的皮肤上是有鳞片的。事实上离片椎类的体表普遍都覆盖着鳞片或骨板。上面说的Melanerpeton tenerum的身体上就布满了1毫米大小的细鳞[8]。

一种二叠纪离片椎类Trimerorhachis(属于双椎蜥亚目Dvinosauria,上回提到过)的单个鳞片印痕。它的鳞片分为上下两层,表面是平行的纵纹(A),下层则是年轮状的生长纹(B),我们甚至可以数出来这条两栖动物活了16岁左右。图片来源自[9]。
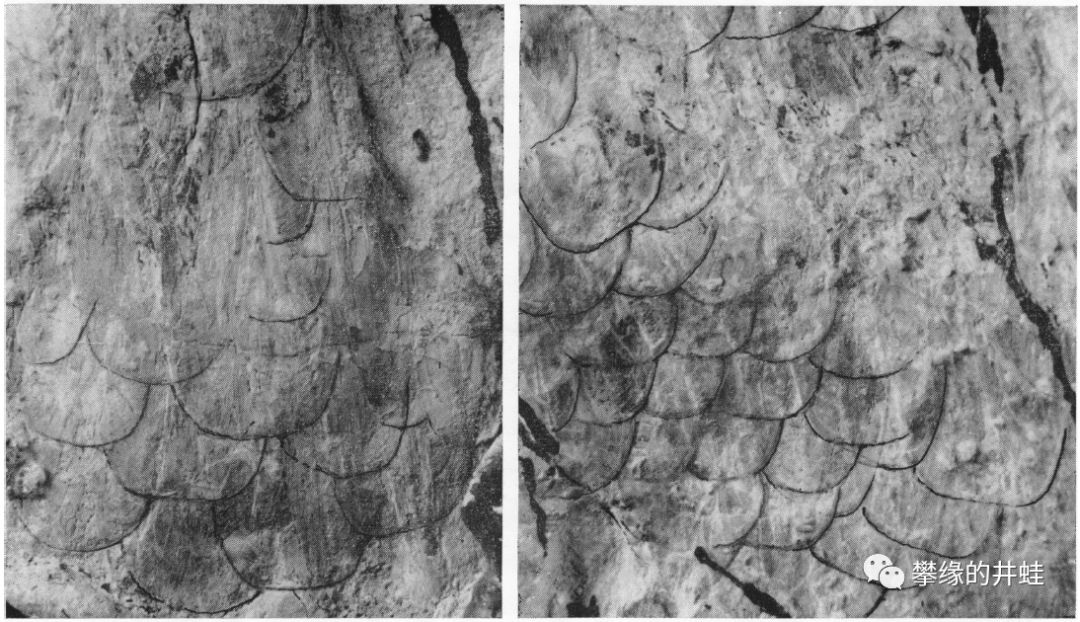
Trimerorhachis化石上的皮肤印痕,它的鳞片互相堆叠覆盖,排列方式很像今天的高等辐鳍鱼类。图片来源自[9]。

Trimerorhachis复原图,模样大小都和一条娃娃鱼差不多。如果它生活到今天,如果被大量养殖作为合法食材,那烹饪的第一个步骤是一定是刮鳞(天呐你对食物,对生命有一丝敬畏之心吗?!)。图片作者:Nobu Tamura。
遗憾的是,保留皮肤结构的化石极其罕有,我们可能永远也不会知道两栖动物是在何时褪去一身鳞甲,变成现在这种滑腻腻黏糊糊的样子。但是,至少从牙齿类型和骨骼结构上看,双疏螈类和滑体两栖类的确关系非常密切。证实离片椎类起源说所缺少的,只有决定性的化石证据——介于二者之间的过渡类型了。
所以当这块化石出现在研究者面前时,可以想象他们的心情是何等的欣喜与激动:
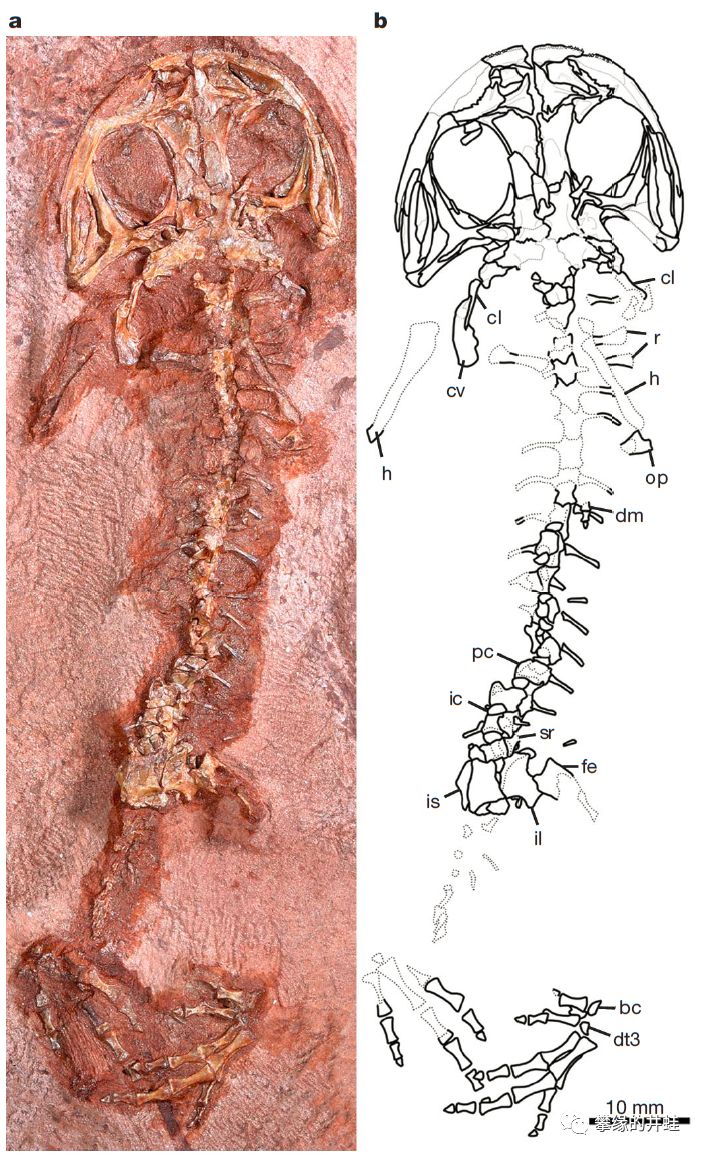
这只动物生活在早二叠世的德克萨斯。它的头骨已经非常接近蛙形类了。但脊柱,肋骨,指骨还保留着许多离片椎类的特征,研究者们把这个梦寐以求的过渡类型命名为:原蛙Gerobatrachus hottoni,但这有点厚此薄彼,因为它也是(或者非常接近)有尾目的祖先。我更喜欢新闻报导中的名字:蛙鲵(Frogamander)。图片来源自[10]。
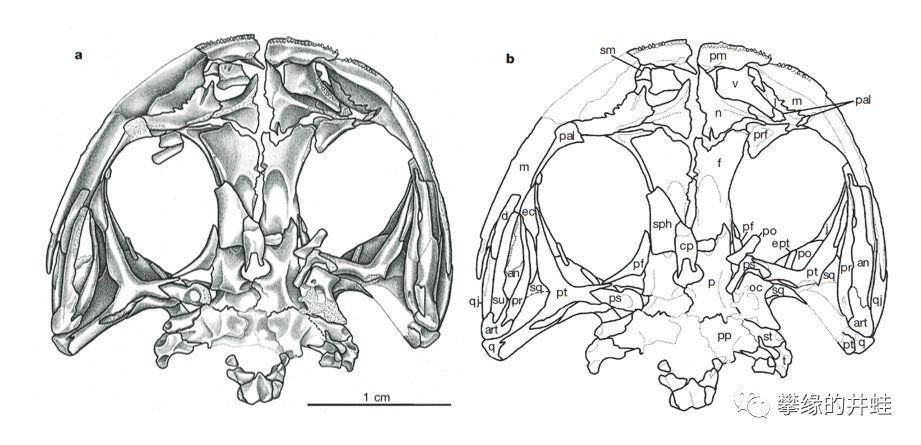
原蛙的头骨结构。一堆小碎骨,两个空荡荡的大眼洞,非常符合蛙形总目的特征。原蛙生活在二叠纪早期,和分子钟推测的无尾目和有尾目的共同祖先的年代相符。它和无足目的共同祖先还要再早一些。图片来源自[10]。
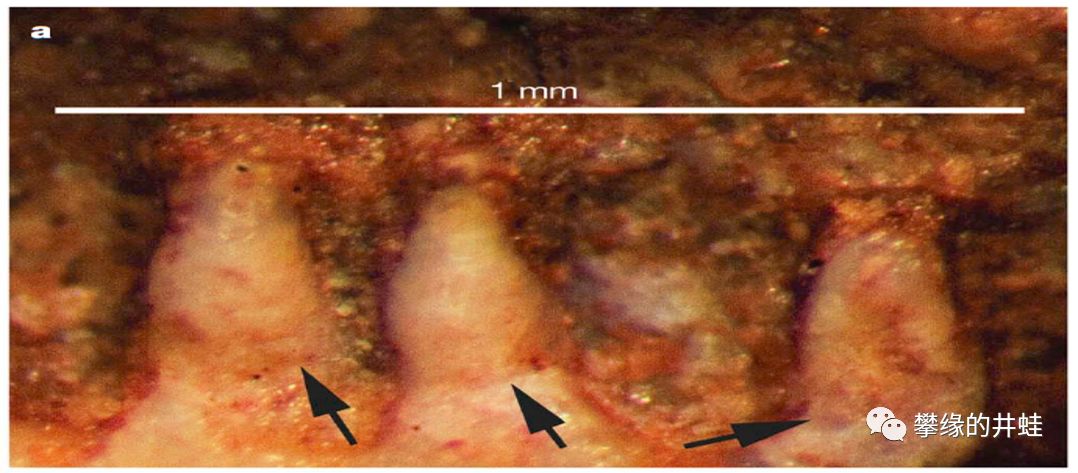
原蛙的牙齿是典型的Pedicellate teeth,箭头标示的是齿冠从管子里伸出来的位置。图片来源自[10]。

原蛙复原图。因为化石缺失了皮肤,尾巴和大部分肢骨,研究者们只能把它想象成一条长着鳞片的“短尾巴蝾螈”。图片来源见水印。
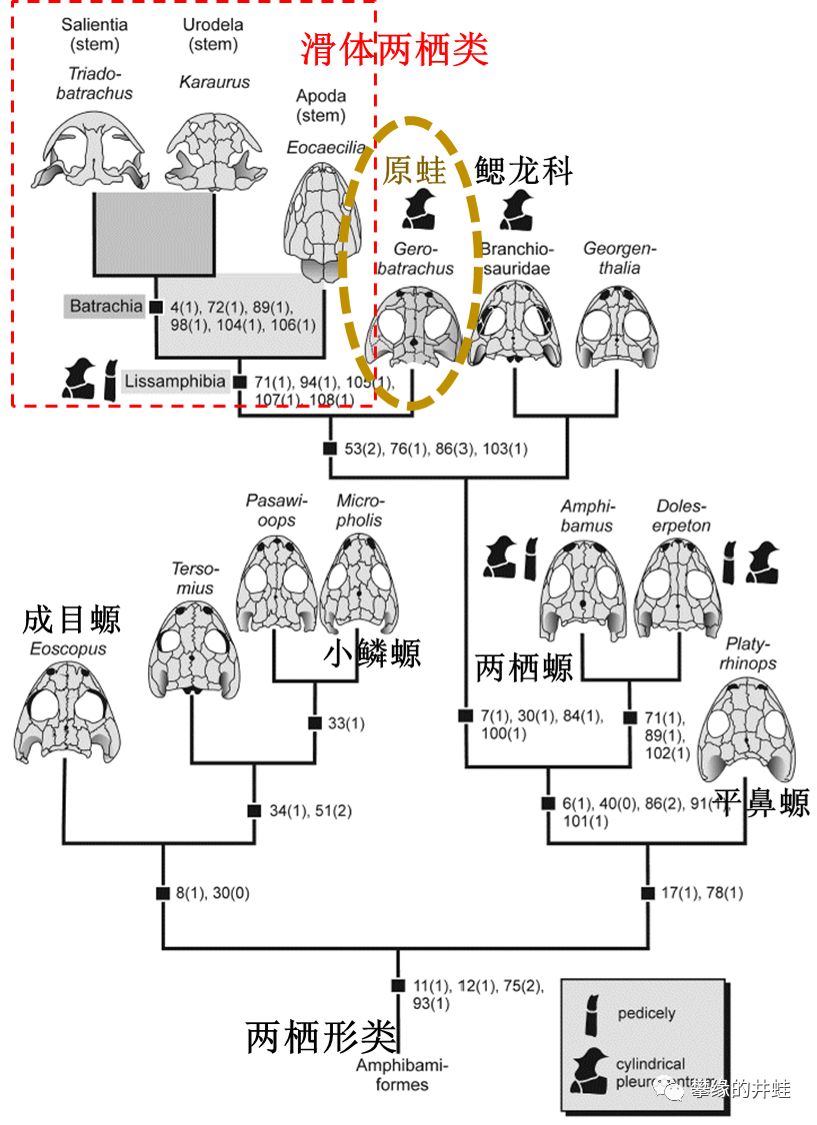
原蛙化石的横空出世,补全了从双疏螈类到滑体两栖类——准确来说是到无尾目和有尾目——的演化链条。无足目可能和双疏螈类关系较远,三叠纪的章回再介绍。现在学界基本上接受了滑体两栖类是离片椎类的直系后裔。好了,快去压棺材板吧。图片来源自[6]。
在分类上,原蛙被归入离片椎类-两栖形类-双疏螈类-两栖螈科(Amphibamidae)-原蛙属。它的一只脚已经踏进了滑体两栖类的圈子。而就在二叠纪刚刚以异常惨烈的方式结束后,真正的滑体两栖类就迫不及待地出现了。
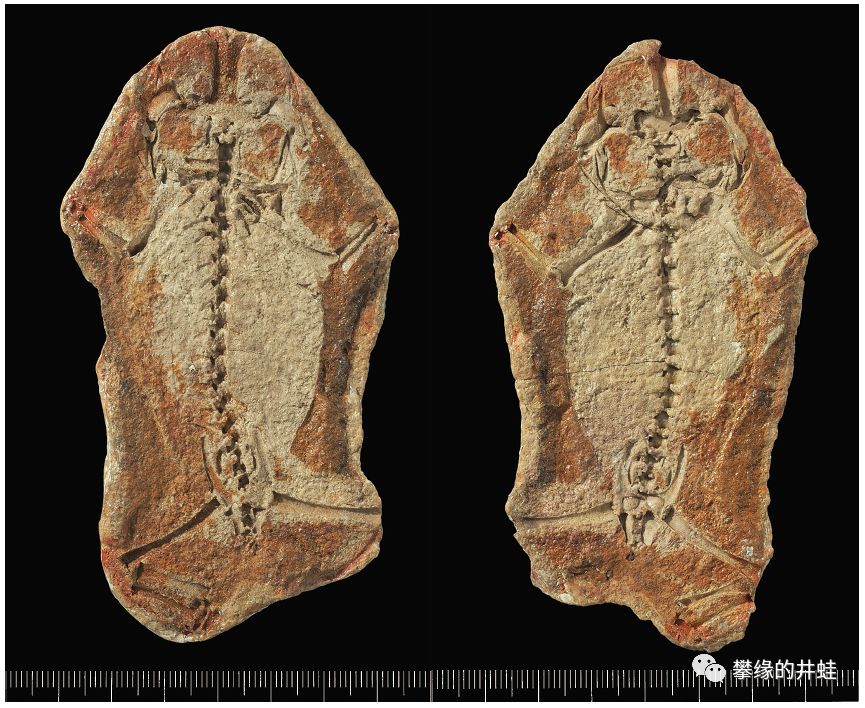
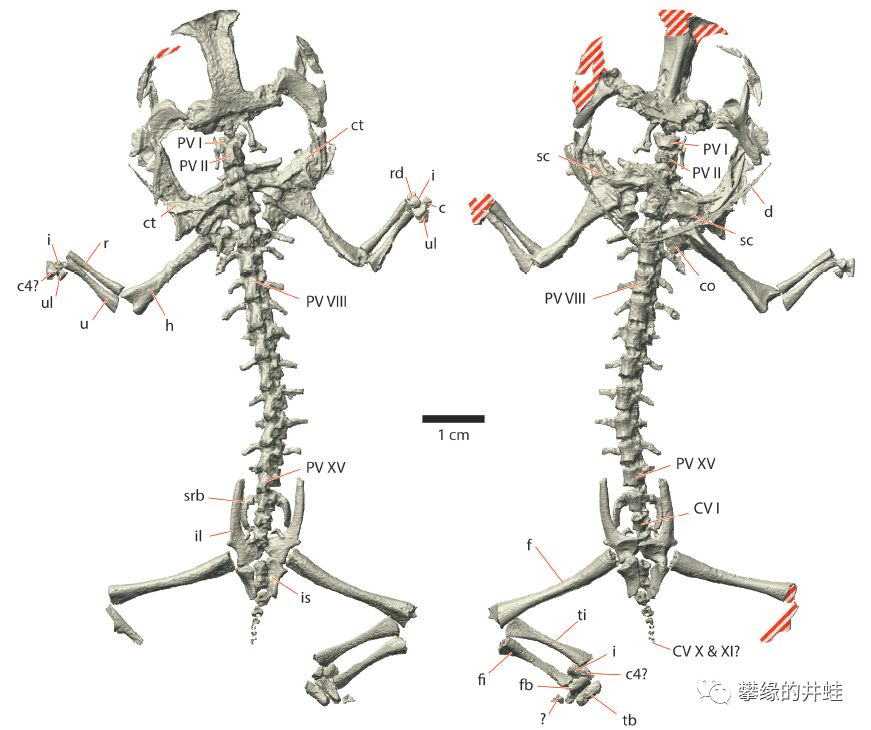
目前发现的最早的滑体两栖类:三叠尾蛙Triadobatrachus massinoti化石和骨骼结构。发现地:马达加斯加。它的屁股后面有拖着一根卖萌用的小尾巴。图片来源自[11]。

三叠尾蛙复原图。它是两栖类演化史上的另一个里程碑,等到三叠纪篇再详细介绍。图片来源自网络。
当古两栖动物终于从淡水霸主的宝座上跌落当巨大的躯体,厚重的头颅,坚硬的皮肤,强健的力量,凶猛的习性,所有的这一切都不能再维系生存时,一个毫不起眼的,光溜溜,软趴趴,滑腻腻的小小旁支,在硬骨鱼类和羊膜动物的夹逼之下,硬是开辟出一些独特的生存空间,顽强地挺过三次大灭绝事件,把离片椎类的血脉延续到今天。无论如何,这都是一项了不起的成就。
所以,我能从棺材板上下来了吗?
地球名片
生物分类:动物界-脊索动物门-脊椎动物亚门-四足形类-滑体两栖类
存在时间:二叠纪早期至现代
现存种类:约6146种(无尾目5420种,有尾目559种,无足目167种)
化石种类:未找到完整统计
生活环境:淡水
代表特征:体型巨大,吻部特化成长嘴
代表类群:各种蛙,蟾蜍,鲵,蝾螈,鱼螈,蚓螈
[1] Danto M, Witzmann F, Fröbisch NB (2016), Vertebral Development in Paleozoic andMesozoic
TetrapodsRevealed by Paleohistological Data. PLoS ONE 11(4): e0152586.doi:10.1371/journal.
pone.0152586
[2] Jason S. Anderson, Focal Review: TheOrigin(s) of Modern Amphibians. Evol Biol (2008), 35: 231–247, DOI10.1007/s11692-008-9044-5
[3] PENG ZHANG, HUI ZHOU, YUE-QIN CHEN, etal., Mitogenomic Perspectives on the Origin and Phylogeny of LivingAmphibians.Syst. Biol. 54(3):391–400, 2005
[4] Anna B. Vassilieva, Sergei V. Smirnov,DEVELOPMENT AND MORPHOLOGY OF THE DENTITION IN THE ASIAN SALAMANDER, Ranodonsibiricus (URODELA: HYNOBIIDAE). Russian Journal of Herpetology, Vol. 8, No. 2,2001, pp. 105–116
[5] TROND SIGURDSEN, JOHN R. BOLT, THELOWER PERMIAN AMPHIBAMID DOLESERPETON (TEMNOSPONDYLI: DISSOROPHOIDEA), THEINTERRELATIONSHIPS OF AMPHIBAMIDS, AND THE ORIGIN OF MODERN AMPHIBIANS. Journalof Vertebrate Paleontology, 30(5): 1360–1377, September 2010
[6] Rainer R. Schoch, The putativelissamphibian stem-group: phylogeny and evolution of the dissorophoidtemnospondyls. Journal of Paleontology, doi: 10.1017/jpa.2018.67
[7] RALF WERNEBURG, TIMELESS DESIGN:COLORED PATTERN OF SKIN IN EARLY PERMIAN BRANCHIOSAURIDS (TEMNOSPONDYLI:DISSOROPHOIDEA). Journal of Vertebrate Paleontology, 27(4): 1047–1050, December2007
[8] Boy, J. A. 1986. Studien über dieBranchiosauridae (Amphibia: Temnospondyli). 1. Neue und wenig bekannte Artenaus dem mitteleuropäischen Rotliegenden (?Oberstes Karbon bis unteres Perm). PaläontologischeZeitschrift 60: 131–166
[9] Edwin H. Colbert, Scales in the PermianAmphibian Trimerorhachis. American Museum Novitates, Number 1740, June 29, 1955
[10] Jason S. Anderson, Robert R. Reisz,Diane Scott, et al., A stem batrachian from the Early Permian of Texas and theorigin of frogs and salamanders. NATURE, Vol 453, 22 May 2008
[11] Eduardo Ascarrunz, Jean-Claude Rage,Pierre Legreneur, et al., Triadobatrachus massinoti, the earliest knownlissamphibian (Vertebrata: Tetrapoda) re-examined by μCT scan, and theevolution of trunk length in batrachians. Contributions to Zoology, 85 (2),201-234 (2016)