
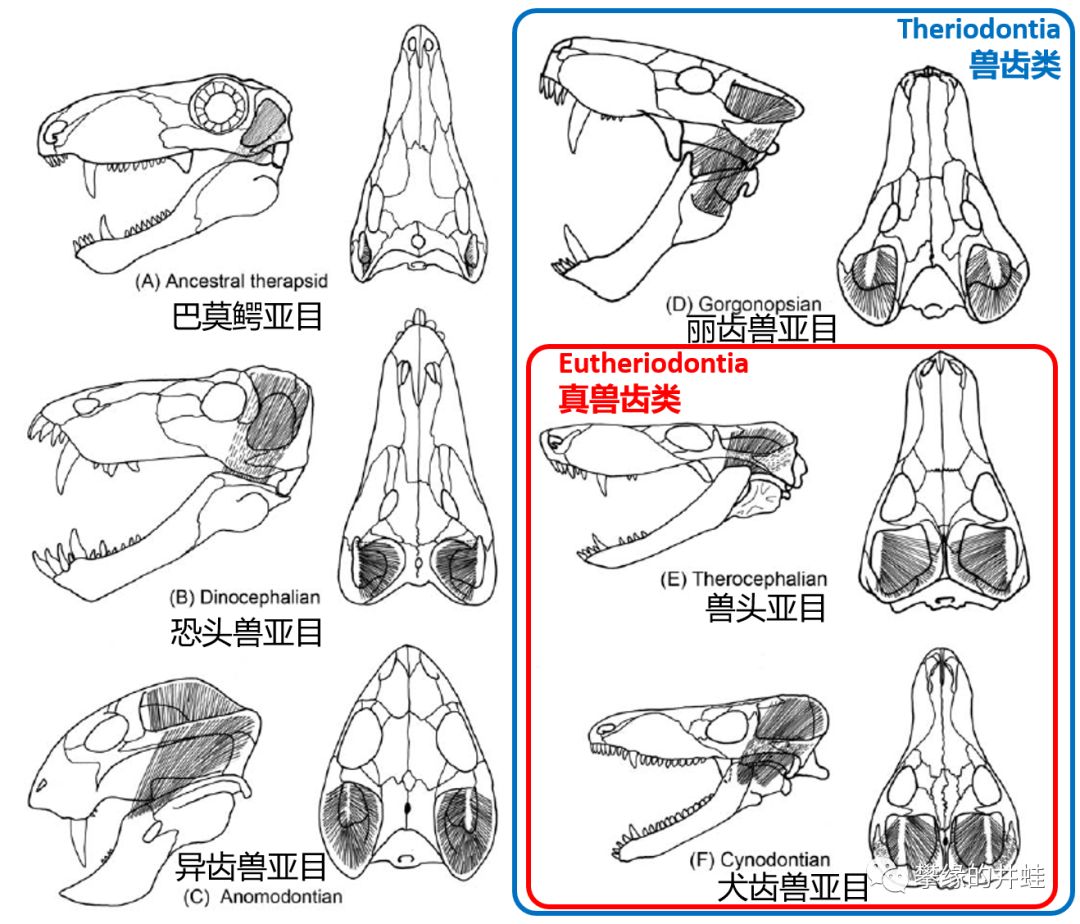
盘龙目在中二叠世全灭,标志着合弓纲的“蜥蜴形”这条后路被彻底堵死了。它们能做的,只有前进前进不择手段地前进,去探寻全新的基本构型和生存模式。
继之而起的兽孔目中,巴莫鳄亚目大体上处在“蜥蜴形”向“兽形”的过渡阶段。演化程度更高的兽孔类大体上出现了三个主要的演化分支:
一个是在中二叠世盛极一时的恐头兽亚目。植食性的貘头兽类,肉食性的安蒂欧兽类还有一堆杂食性的乱七八糟兽类,掀起了一波多样性的高潮,占据的基本上就是后来(非鸟)恐龙的生态位(就连名字里也都带个“恐”:Dinocephalia 和Dinosaur )。恐头兽亚目这一场短暂的盛衰枯荣倒有点像是恐龙王朝命运的一次预演,二者的退场也都是一样的猝然而彻底。
恐头兽类凋零后,空出的植食性生态位主要被异齿兽亚目,尤其是二齿兽次亚目填补。这个极其特化的类群代表着兽孔目的另一个尝试方向。二齿兽类放弃了灵活的身姿,专心去当大大小小的移动发酵罐。为此不惜用硬质喙取代了合弓纲的祖传大杀器异型齿。当那些小型种类在二叠纪末大灭绝中损失殆尽后,剩下一班粗笨夯货们迟早会陷入万劫不复的境地。
第三个演化支被称为“兽齿类(Theriodontia )”,包括丽齿兽亚目,兽头亚目(Therocephalia )和犬齿兽亚目(Cynodontia )。其中兽头亚目和犬齿兽亚目亲缘关系很近,合称为“真兽齿类(Eutheriodontia )”。兽齿类的演化路线和二齿兽类几乎完全相反:强化运动和感知能力,牙齿进一步分化,配合强大的咬合肌群,这些特质使得兽齿类接替肉食性恐头兽,成为二叠纪最主要的陆生掠食者。

兽孔目主要类群代表种类的头骨,牙齿以及咬合肌结构复原图。A:巴莫鳄Biarmosuchus;B:巨型兽Titanophoneus;C:始二齿兽Eodicynodon;D:熊颌兽Arctognathus;E:Olivierosuchus;F:原犬鳄龙Procynosuchus。图片来源自[1]。
从上图中不难看到,相比丽齿兽亚目,真兽齿类(兽头亚目和犬齿兽亚目)牙齿形态分化程度更高,下颌更加接近哺乳类的单一齿骨结构,颞颥孔更大,齿骨- 鳞骨关节初步成型。
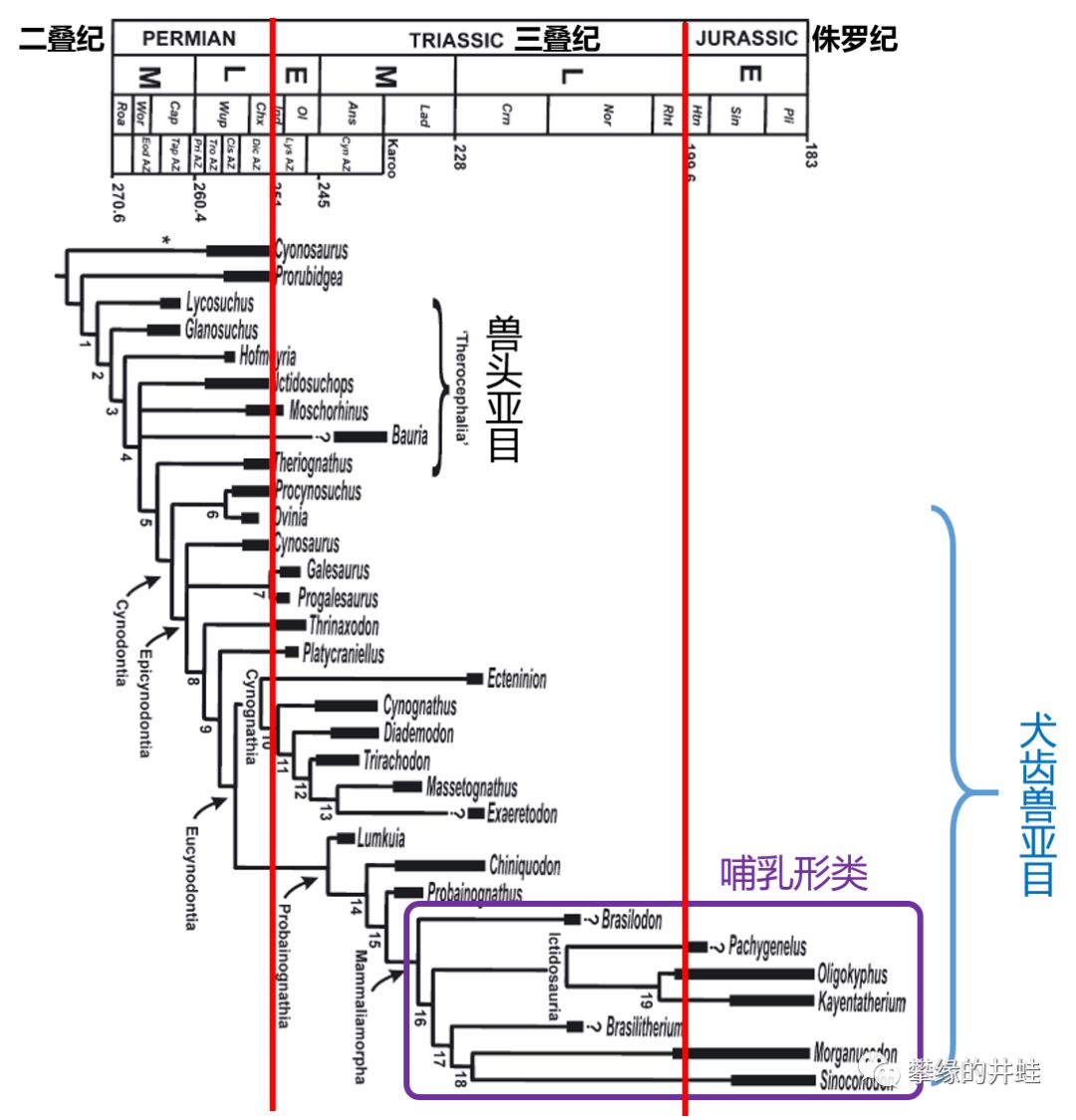
真兽齿类演化谱系和生存时代简图。它们是兽孔目中和哺乳类亲缘关系最密切的类群。包括哺乳动物在内的“哺乳形类(Mammaliamorpha)”其实是犬齿兽亚目在中生代后期仅存的一个演化支。与其说兽头兽类和犬齿兽类“像”哺乳动物,其实是哺乳动物继承并光大了真兽齿类的结构特征。图片来源自[2]。
因为这种“亲密关系”,人们在复原兽头兽和犬齿兽时总会自觉不自觉地赋予大量哺乳动物的外形特征,结果复原图看上去基本上就是“状如小狗而很凶猛”的样子。
比如这样的:
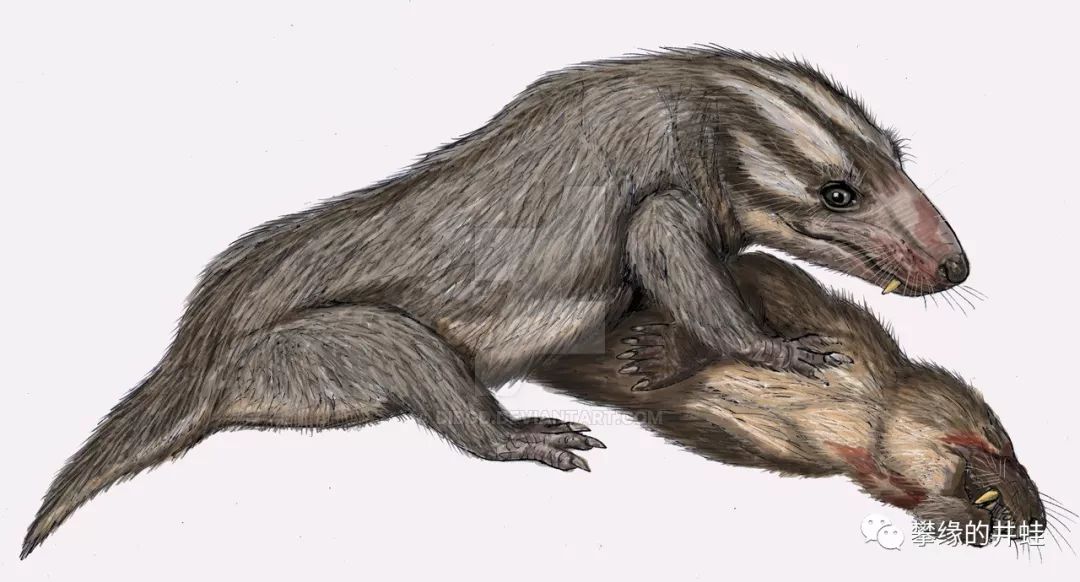
Moschowhaits iavjuschkovi(兽头亚目)生活在二叠纪末,化石产自俄罗斯和东欧地区,是一种中大型肉食动物(头骨长25厘米)。被捕食的是一种植食性的兽头兽Purlovia maxima。图片来源自[3]。

另一种俄罗斯出土的二叠纪晚期兽头兽Karenites ornamentatus。图片来源自网络。

Gorynychus masyutinae是俄罗斯发现的中二叠世兽头兽。体型和狼相当,拥有发达的犬齿。图片来源自[4]。

德韦纳兽Dvinia prama头骨化石和复原模型。属于早期犬齿兽类,在俄罗斯发现,生存于晚二叠世。头骨长约10厘米,可能主要捕食昆虫和小型脊椎动物。图片来源自网络。

德韦纳兽(左)和同一化石群发现的中型二齿兽Fortunodont rautscholdi(右)体型对比。图片来源见水印。
总感觉,怎么说呢,狗头鼠脸的,辨识度不高。

当然也有比较精奇的复原,比如这个“原犬鳄龙Procynosuchus(犬齿兽亚目)”。狗头也太逼真了也有点受不了,而且好歹加上点儿毛吧。图片来源自网络。

原犬鳄龙骨架复原。按照惯例“Procynosuchus”应该译成“原犬鳄兽”的,但井蛙看到的所有中文资料都用“原犬鳄龙”,就不擅改了。原犬鳄龙是犬齿兽的早期种类,从骨骼结构来看,头骨后面的部分和巴莫鳄类并没有多大区别。图片来源自网络。
复原形象都这样,那没皮没脸没肉没毛的头骨更是要为难死脸盲患者了。
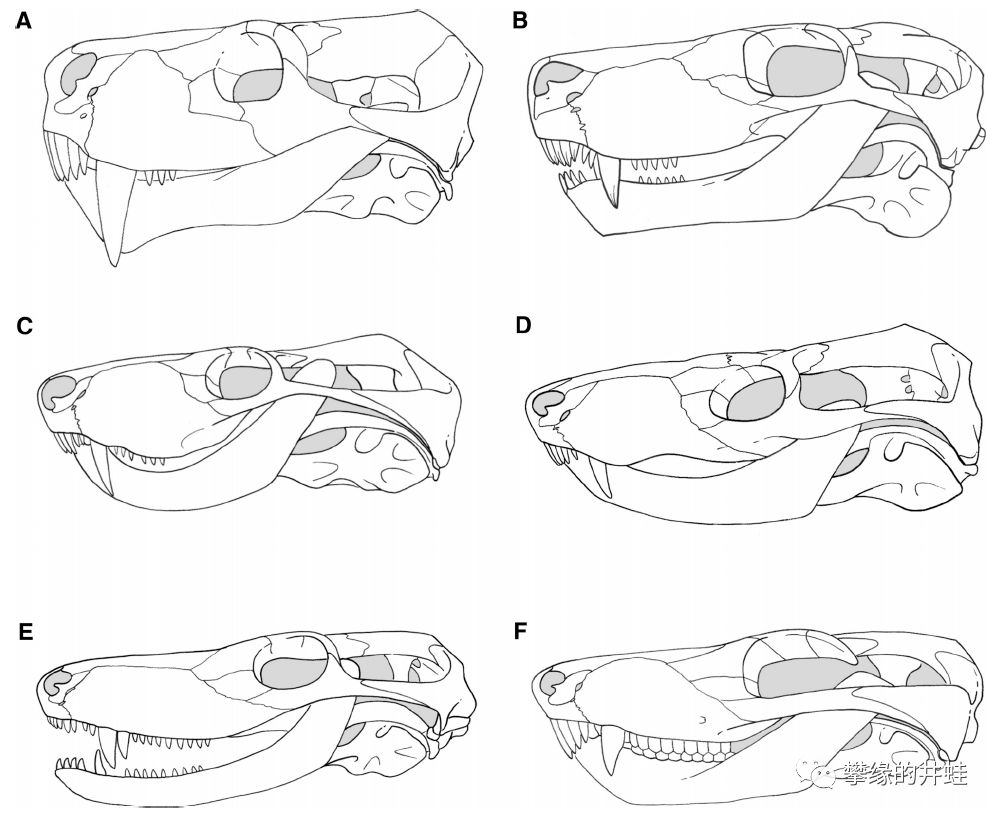
兽头亚目主要类群代表物种的头骨结构。A:麝喙兽Moschorhinus kitchingi(弱颌兽科);B:Promoschorhynchus(弱颌兽科);C:Mirotenthes digitipes(霍夫梅里兽科);D:兽颌兽Theriognathus microps(Whaitsiidae);E:似鼬鳄兽Ictidosuchoides longiceps(包氏兽科);F:包氏兽Bauria cynops(包氏兽科)。它们的齿骨末端以及上翘插入鳞骨,但隅骨和上隅骨还没完全退化。图片来源自[5]。
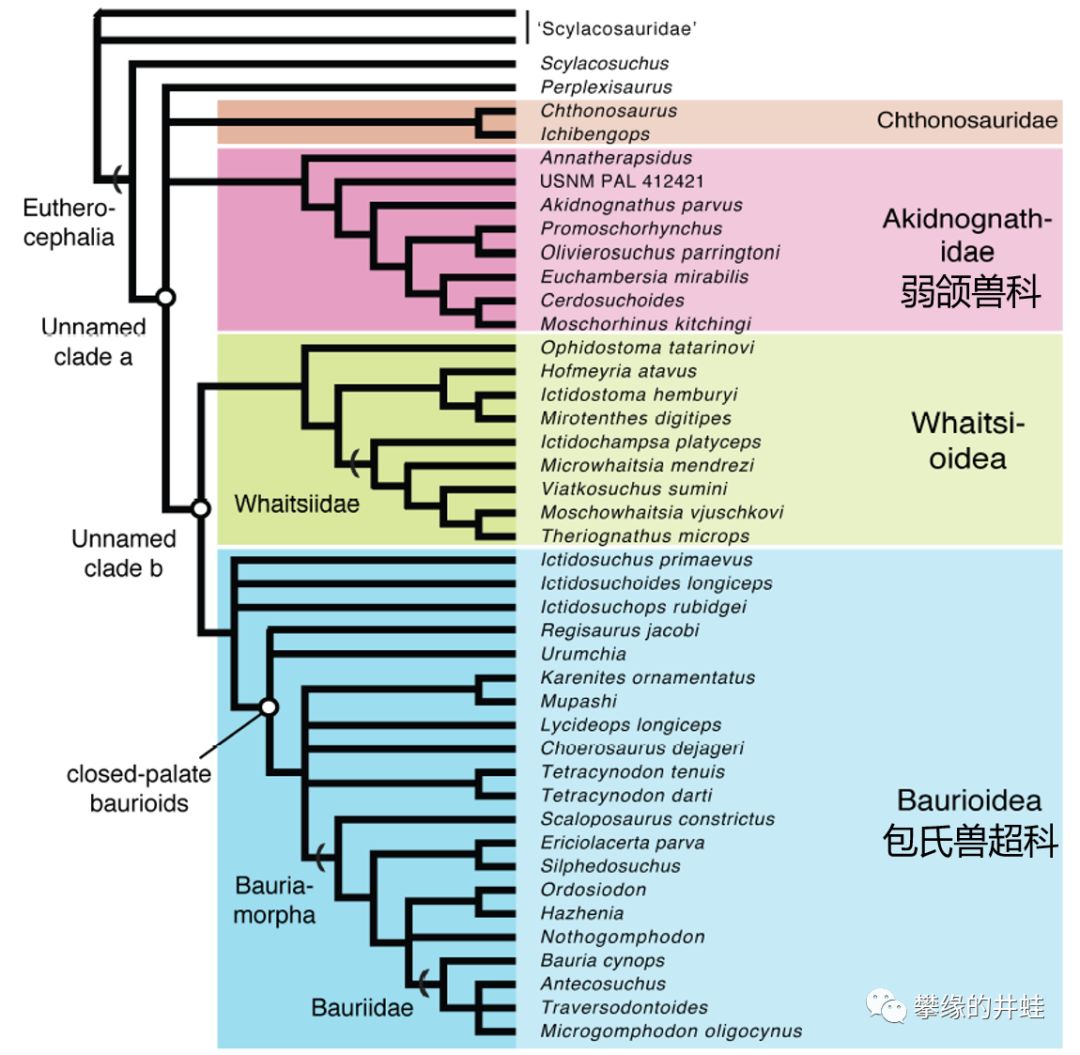
兽头亚目的演化谱系简图。可以对应上面的头骨形态来看,包氏兽大多植食性,而其他类群基本都是肉食性的,它们的牙齿形态自然有很大差异。图片来源自[6]。
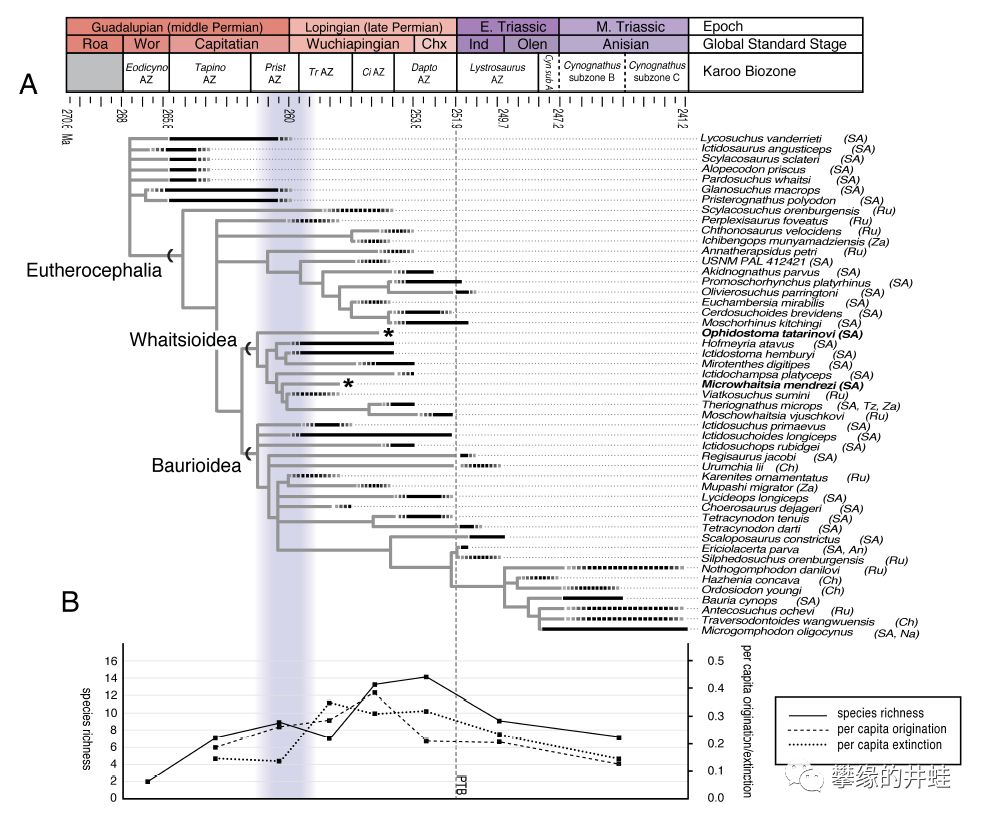
兽头亚目主要类群代表物种的生存年代。二叠纪末大灭绝沉重打击了兽头兽类的多样性,除了少数包氏兽外几乎全灭。图中括号里也标出了各种兽头兽的发现地:An-南极洲,Ch-中国,Na-纳米比亚,Ru-俄罗斯,SA-南非,Tz-坦桑尼亚,Za-赞比亚。这么一看,南非真是二叠纪陆羊膜类化石的宝库。图片来源自[6]。
其实兽头亚目在二叠纪家族兴旺。除了类似“狐狼狸獾貂鼬”这样的中小型食肉动物之外,也拓展到了其他的生态位,出现了一些很有特色的种类。
巨型掠食者 Megawhaitsia patrichae 。看名字就知道它和 Moschowhaitsia vjuschkovi 一样,属于Whaitsiidae 。这个科和包氏兽科是二叠纪晚期最重要的两支兽头兽。

Megawhaitsia patrichae生活在二叠纪末的俄罗斯东部,有着兽头亚目种少见的巨大体型。它和大型丽齿兽一样,是同时代二齿兽和锯齿龙的梦魇。图片来源自[3]。
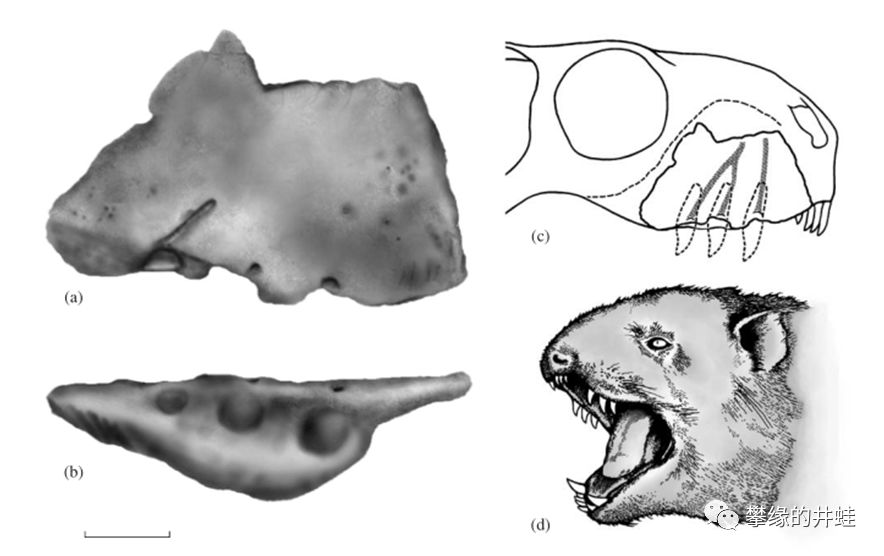
其实Megawhaitsia patrichae只发现了一块右上颌骨的碎片,它的大小和外形是参照其他兽头兽按比例推测出来的。图片来源自[7]。
毒兽 Euchambersiamirabilis 。在BBC 的《Walking with Monsters 》中出场过,编剧安排它用毒液快速猎杀水龙兽。根据新的化石证据,这个设定还是挺靠谱的。

《Walking with Monsters》中,在夜色掩护下伏击迁徙中的水龙兽的Euchambersia mirabilis。
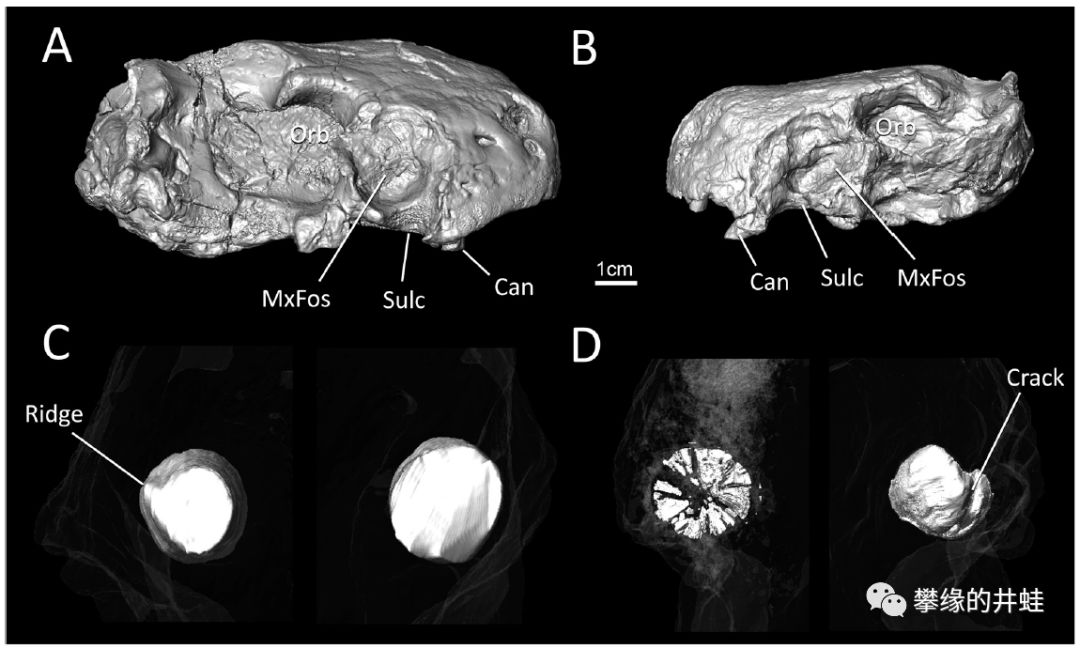
Euchambersia mirabilis会使用毒液狩猎的推测主要源自化石(A和B是目前发现的两个比较完整的头骨标本)上的两个特征。一是犬齿上方的上颌骨有一个很大的凹陷(犬齿窝Maxillaryfossa,上图中MxFos标示的位置),可能用来容纳分泌和储藏毒液的腺体。二是犬齿边缘有粗糙的棱脊(上图中Ridge标示),可能用来撕裂创口,让毒液更多更快地侵入猎物体内。图片来源自[8]。
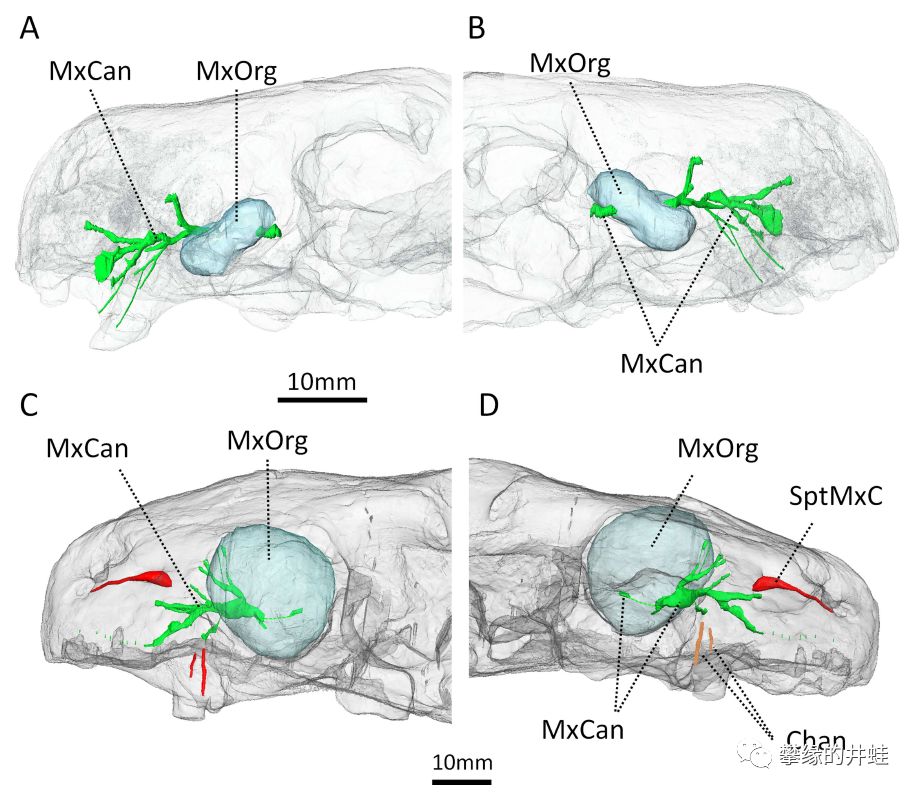
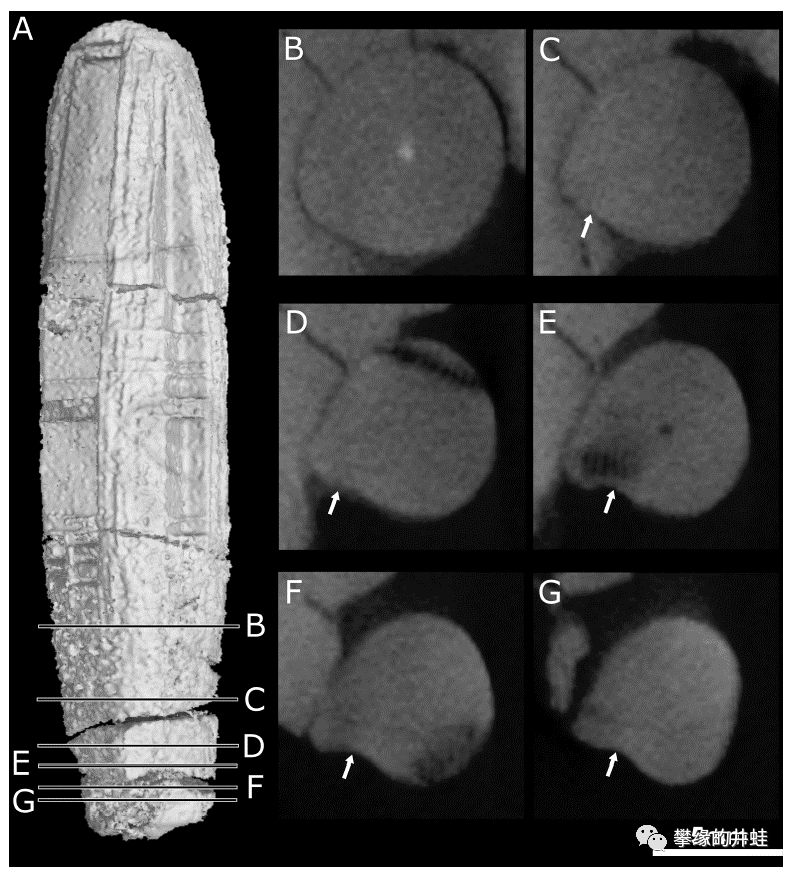
Euchambersia mirabilis头骨化石CT扫描成像。淡蓝色的MxOrg是犬齿窝中的器官,很可能是毒囊。绿色的MxCan是神经系统的痕迹。注意在C,D图中,研究者发现了上颌骨存在通往犬齿根部的细小管道Chan。这个发现更加提高了毒腺存在的可信度。推测毒液就是通过这些管路流入口腔,浸润犬齿,随后进入猎物体内。图片来源自[9]。
Euchambersia mirabilis犬齿的外形和截面,箭头标示了棱脊。犬齿不是中空的,不能像注射器那样直接注入毒液,很可能像现代毒蜥一样,毒液与唾液混合后,从猎物的伤口流入。至于毒液成分是什么,毒性是否剧烈,就不得而知了。图片来源自[9]。
Choerosaurus dejageri ,这个好像还没有中文译名,井蛙就叫它“四锤兽”吧。它的上颌前端,和下颌末端各长着一对骨瘤,这种奇怪的构造在合弓纲中也算独一无二了。
“四锤兽”复原图。外形什么的反正不会太准确,不用细究,注意嘴上的肿包就行了。图片来源自网络。
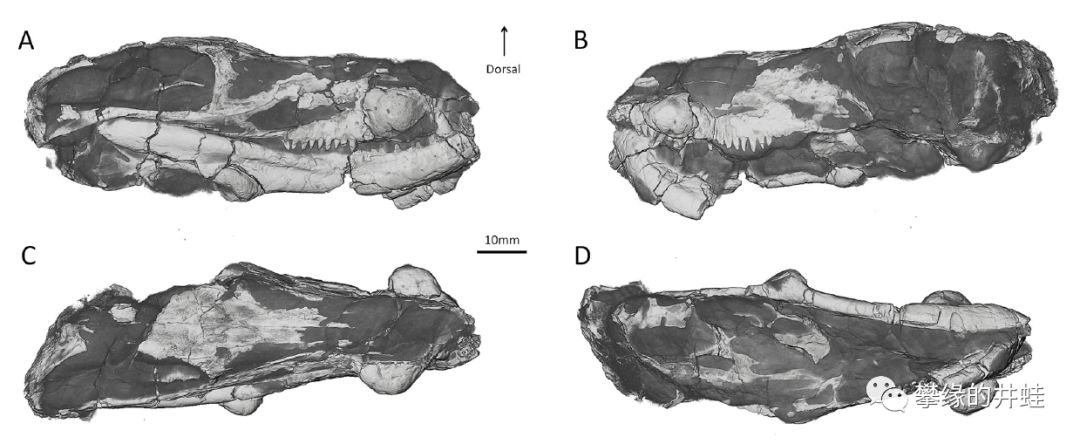
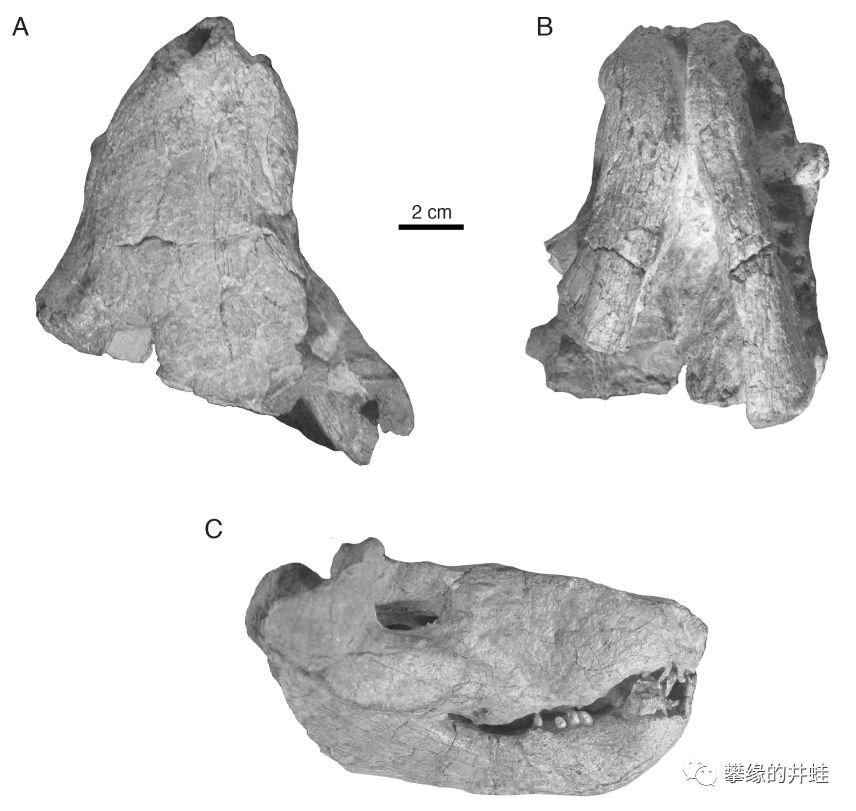
“四锤兽”生活在晚二叠世的南非。这是它的头骨化石,注意骨瘤(C,D图最明显)的位置。图片来源自[10]。
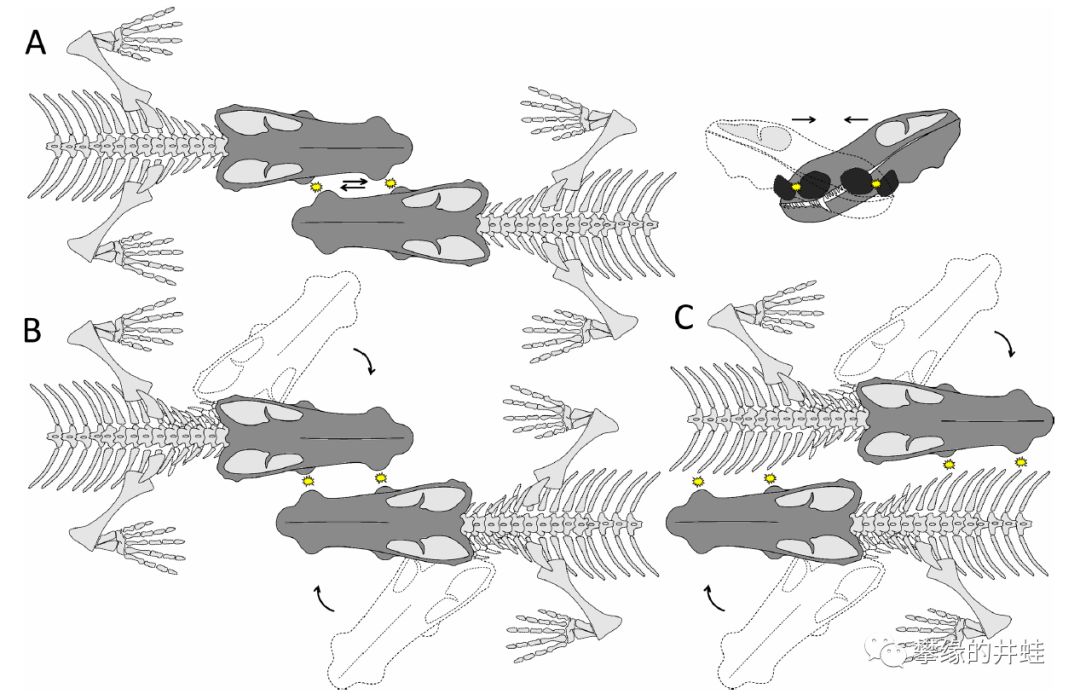
研究者推测雄性“四锤兽”可能用这些骨锤互相撞击,决出各自的地位。这种行为在二叠纪的真兽齿类中非常少见(恐头兽中倒是很常见)。想想两只状如小狗的动物噼里啪啦地轮着脑袋乱撞,还挺好玩呢。图片来源自[10]。
其他的二叠纪兽头兽就比较“大众脸”了。下面列举了一些,大家随便看看吧。
前边说了“Megawhaitsia”,再来个“Microwhaitsia”——Microwhaitsia mendrezi,化石是在南非发现的。图片来源自[6]。

Microwhaitsia mendrezi的体型。虽然名字里有“Micro”,但也不算小呢。图片来源自网络。
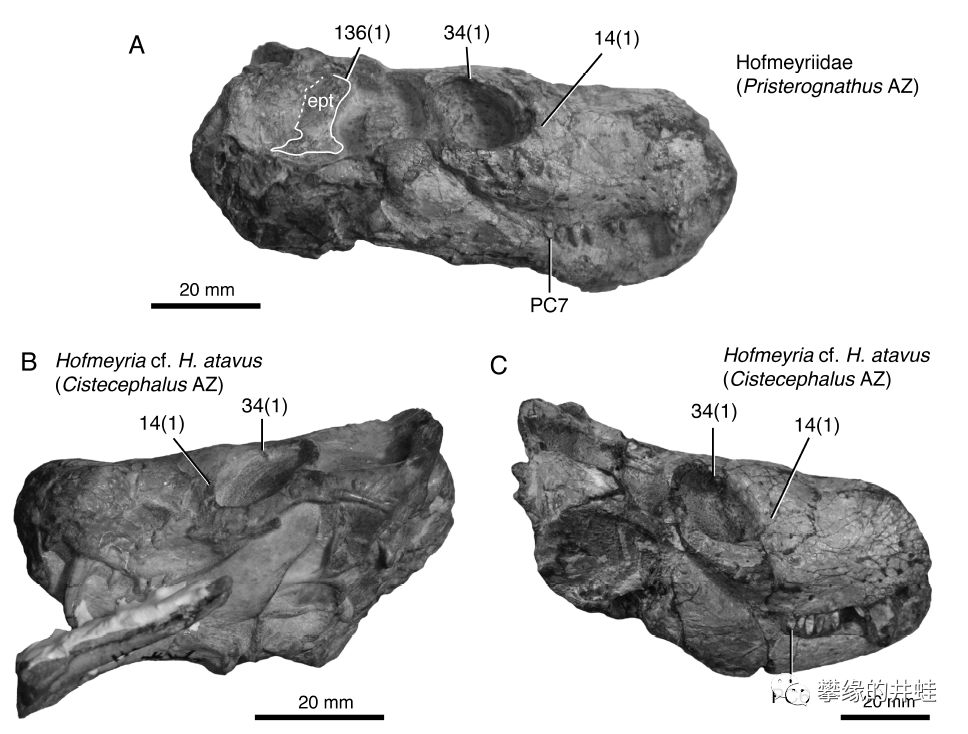
还是南非发现的,两种霍夫梅里兽科的兽头兽。图片来源自[6] 。
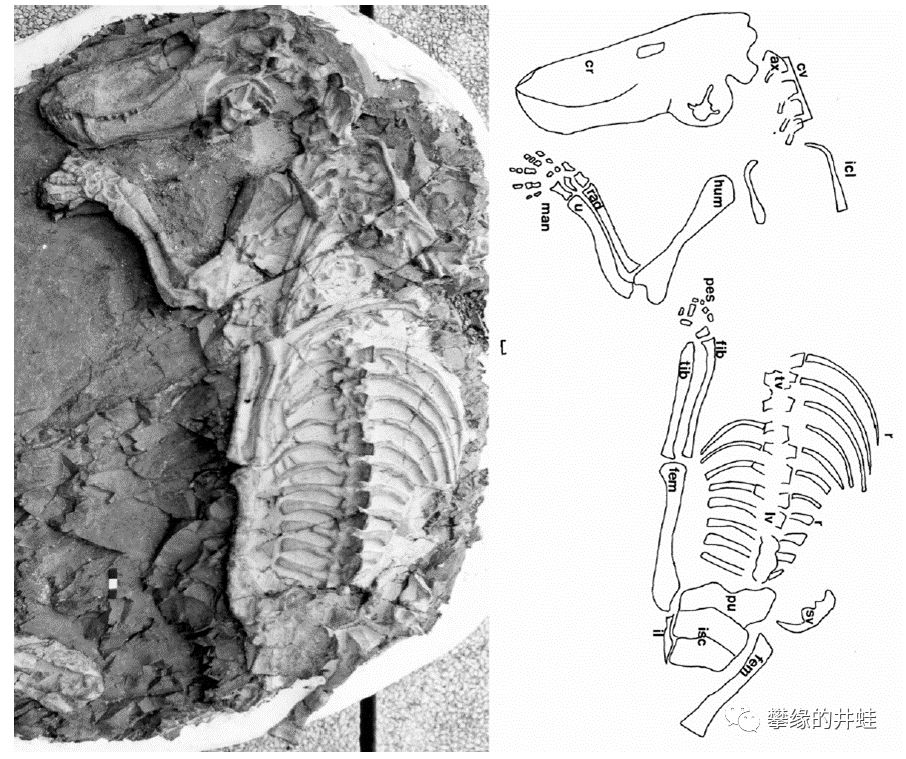
来一个完整度比较高的:又是南非发现的,Glanosuchusmacrops,属于犬齿兽的基干形类。图片来源自[11],标尺长度1厘米(这个标尺我找了半天)。
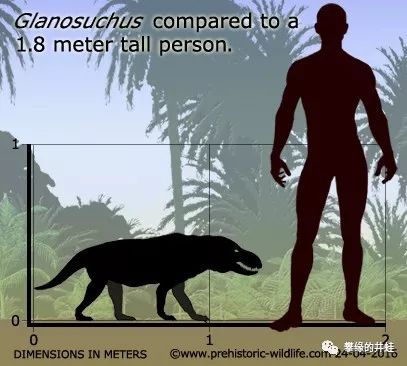
Glanosuchus的体型。根据鼻腔结构,研究者认为它可能是最早演化出恒温(准确地说是内温Endothermy)能力的兽孔类[12]。图片来源见水印。


当然更多的是化石比较零碎的。这是俄罗斯东部发现的中二叠世兽头兽Julognathus crudeli,基本上就拼出来半个脑袋。图片来源自[13],复原图标尺长度2厘米。

Julognathus crudeli复原图。图片来源自网络。
对非专业的读者来说,兽头亚目和犬齿兽亚目都是一副“狗头鼠脸”。其实仔细观察位于口腔内部,犬齿后面的牙齿(犬后齿Postcaninetooth )形态的话,还是比较好区分的。
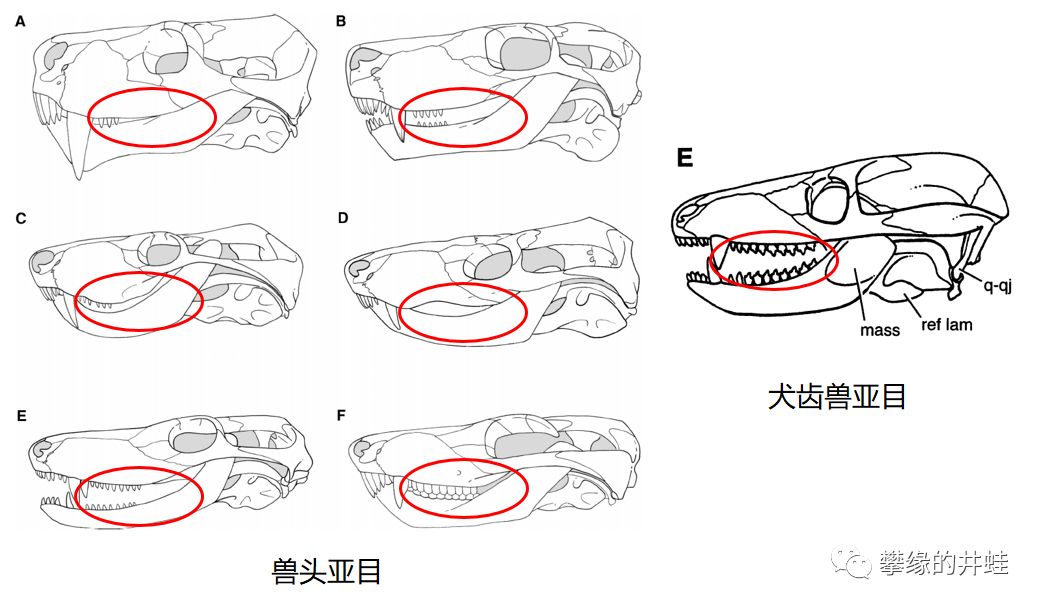
兽头亚目和犬齿兽亚目(原犬鳄龙)头骨对比,注意红圈中的犬后齿。显然犬齿兽类的犬后齿形态更复杂,功能更强大,这就是哺乳动物的臼齿的雏形。图片来源自[5][14]。
二叠纪的犬齿兽亚目种类不多。除了原犬鳄龙和德维纳兽之外,比较重要的也就是凹颌兽( Charassognathus ),犬龙兽( Cynosaurus )和矮貂龙兽( Nanictosaurus )。
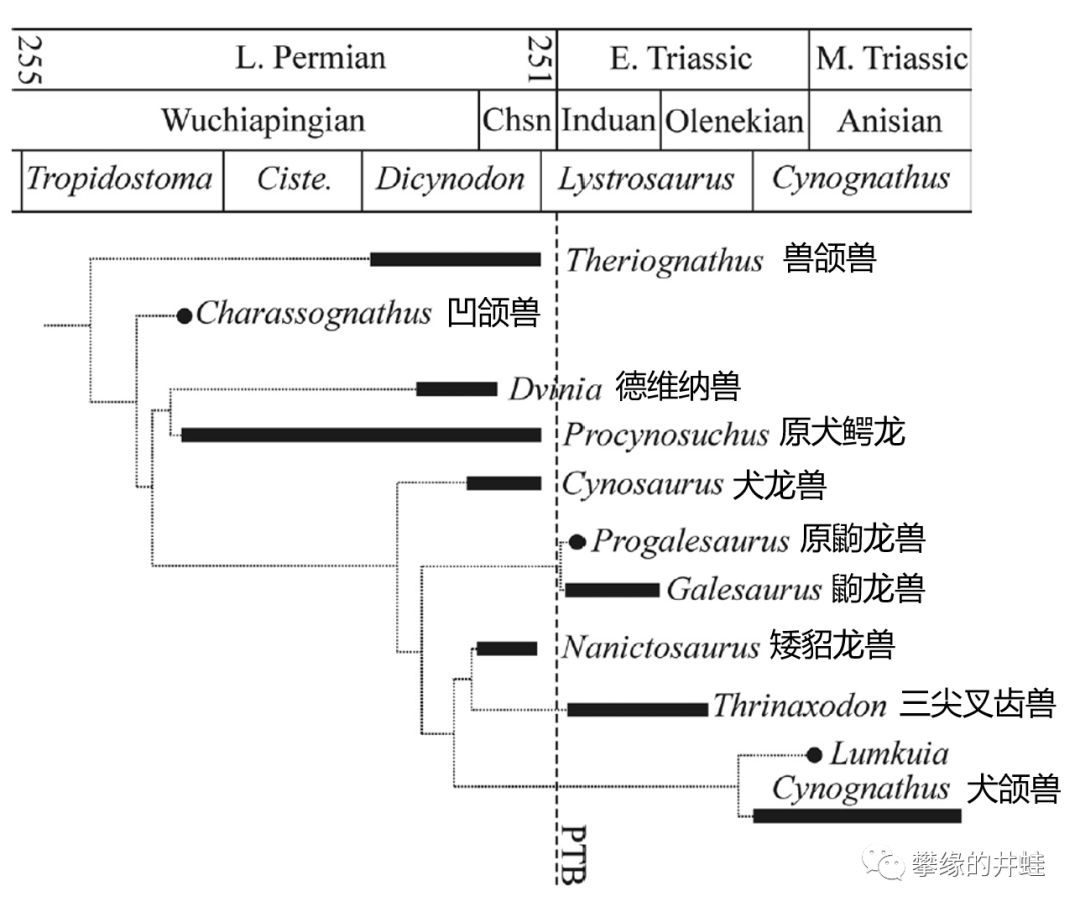
主要早期犬齿兽类演化谱系和生存年代。虚线PTB是二叠纪-三叠纪交界(Permian-Triassic Boundary)。图片来源自[15]。
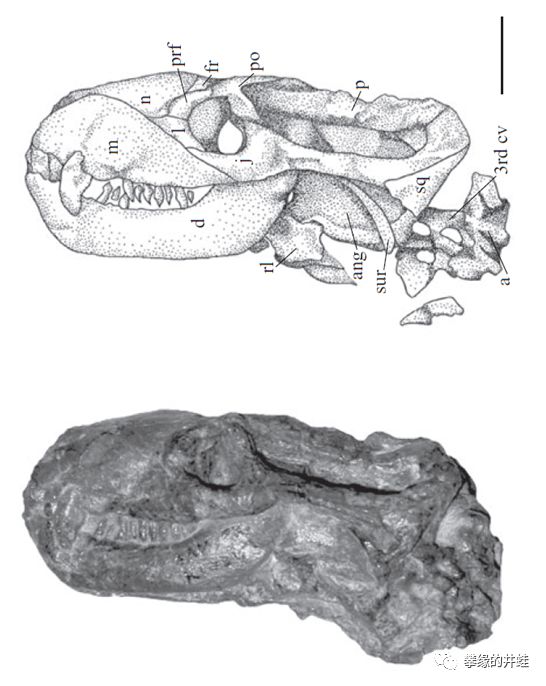
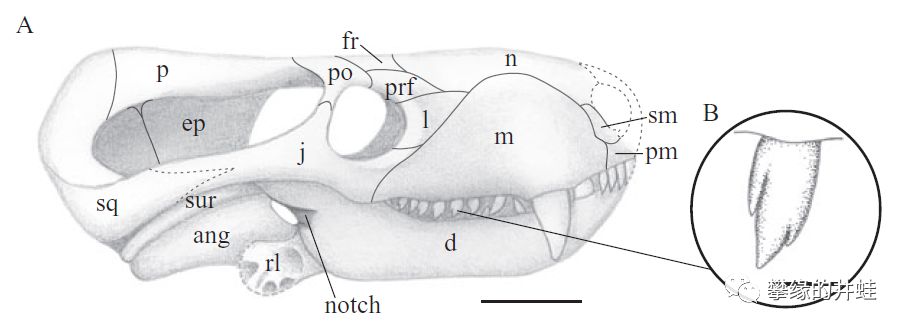
南非出土的凹颌兽Charassognathus gracilis是已发现的最早的犬齿兽类。它的犬后齿已经分裂出多个齿尖,表明此时犬齿兽类已经与兽头兽类分道扬镳。复原图头骨标尺1厘米,犬后齿标尺1毫米。
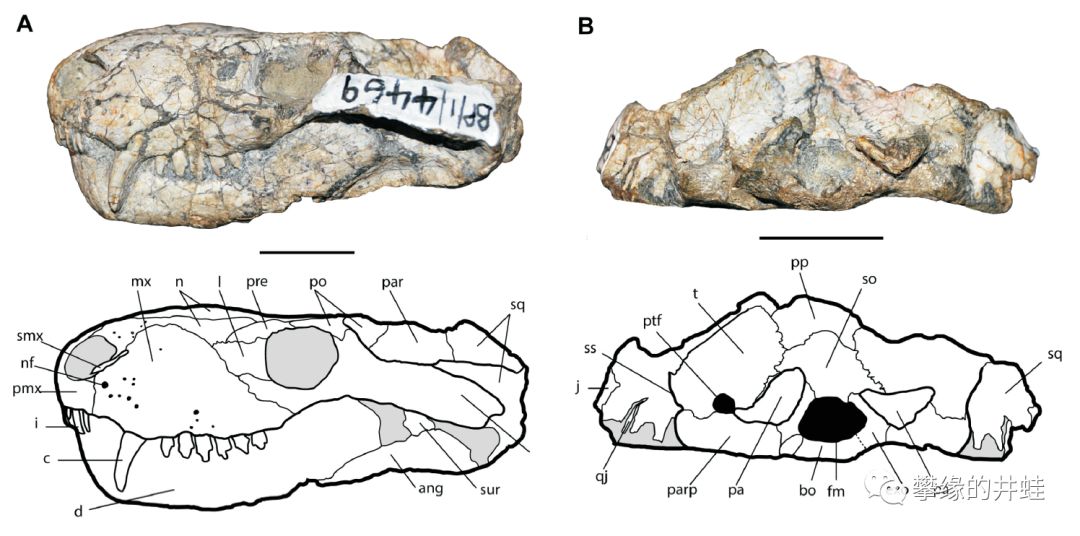

南非发现的犬龙兽Cynosaurus suppostus化石。还是只看犬后齿就行了。图片来源自[17],标尺长度1厘米。
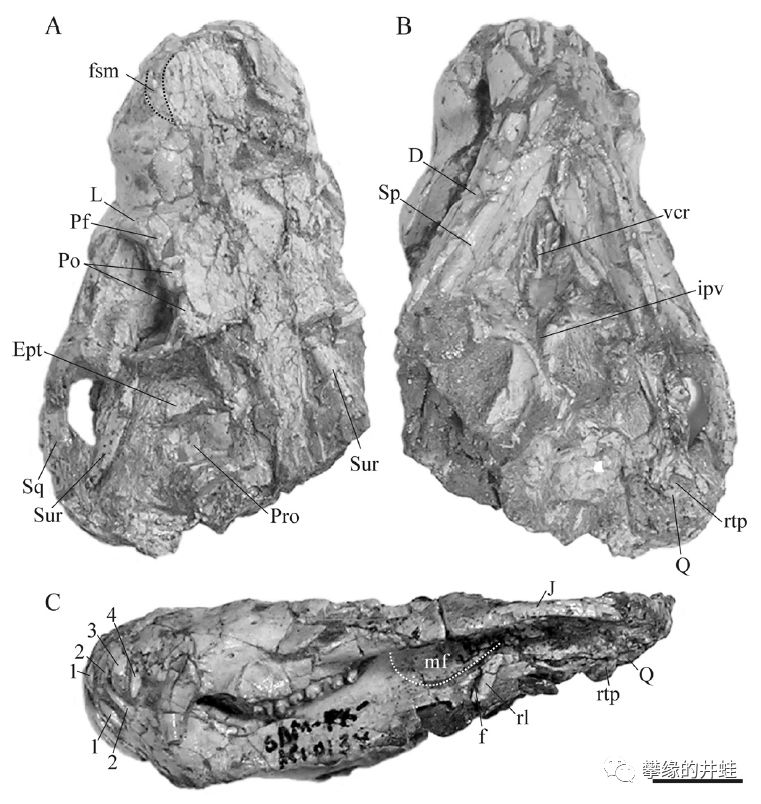
南非新发现的一种二叠纪犬齿兽类,尚未命名。可以清楚地看到犬后齿上像小花瓣似的分裂齿冠,有点像我们的后槽牙了。图片来源自[15],标尺长度1厘米。
当然兽头兽类的犬后齿上也可能出现一些特化,但不会像犬齿兽这样分出多个齿尖,形状这么复杂。

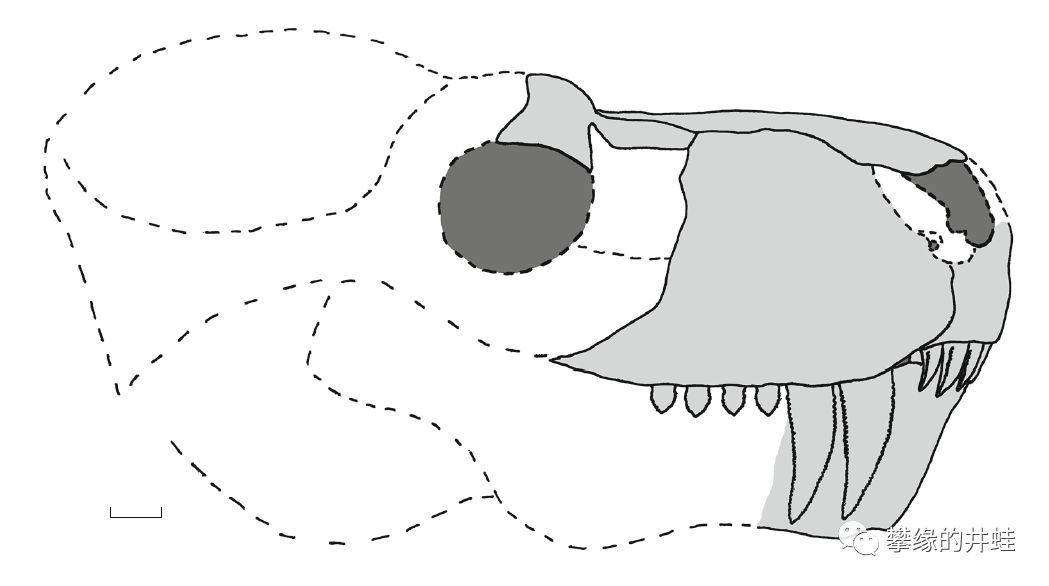
对比一下吧,这是俄罗斯发现的兽头兽Gorynychus sundyrensis(前面有它的近亲Gorynychus masyutinae的复原图)。显然它也在努力改良犬后齿的结构,比如边缘加上了锯齿和沟槽,但和犬齿兽一比,就差得远了。图片来源自[18],复原图标尺长度2厘米。
高度特化的犬后齿让犬齿兽类拥有更强的取食能力。它们可以把食物充分咀嚼,提高消化效率。也可以捡拾丽齿兽,兽头兽丢弃的猎物遗骸,从骨头上啃下紧贴着的肉丝,软骨和韧带。甚至咬碎骨骼,吸食骨髓。凭借这些优势,犬齿兽类熬过了二叠纪末大灭绝,又在三叠纪中期取代了回光返照的兽头兽类。然而面对合弓纲大厦倾颓,蜥形纲迅猛崛起的运势,等待犬齿兽类的,注定是中生代的黯淡岁月。
三叠纪还会有一回介绍兽头兽和犬齿兽之后的故事。许多朋友好奇二叠纪的兽孔类到底有多像或者多不像哺乳动物,井蛙也查了一些资料,下面简单回答几个热门问题。
Q1:二叠纪的兽孔类有毛吗?
A1:从已经发现的化石证据来看,至少一部分二叠纪兽孔类应该是长着毛的。遗憾的是,关于毛发出现和早期演化的化石资料异常稀缺。
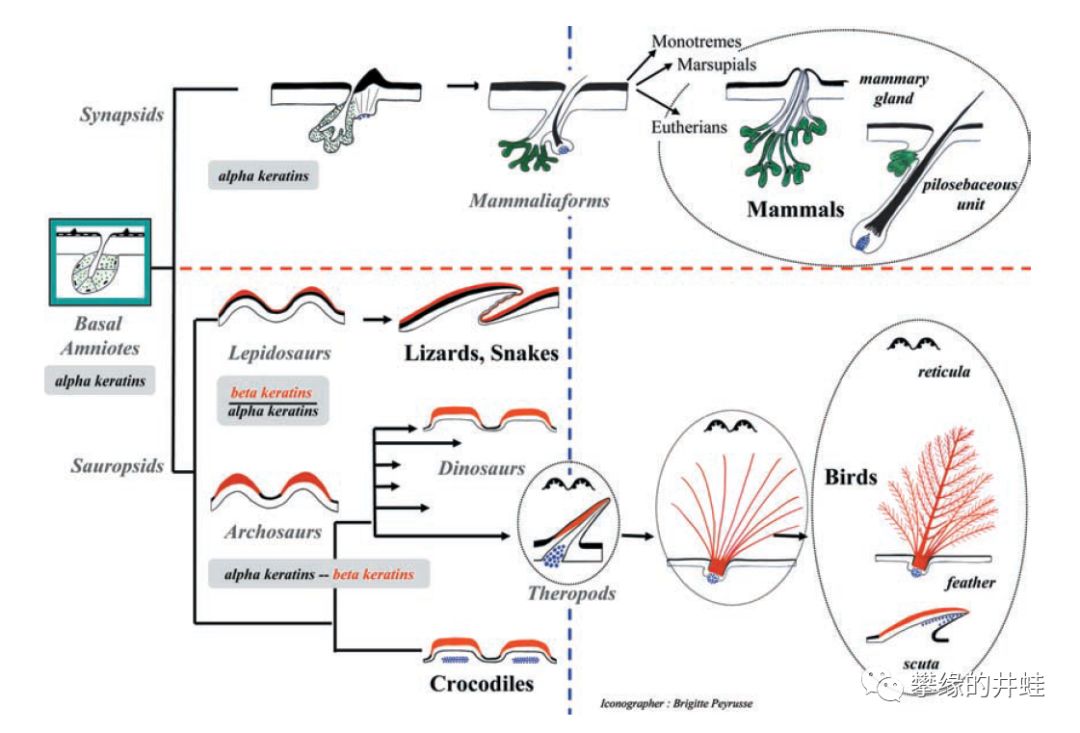
关于毛发(Hair),鳞片(Scale)和羽毛(Feather)演化过程的一种理论。和蜥形纲的羽毛相比,合弓纲的毛发简直粗陋寒酸;而且羽毛的出现和演化过程有着丰富而清晰的化石链条,而毛发就只有一点儿模糊的间接证据。图片来源自[19]。
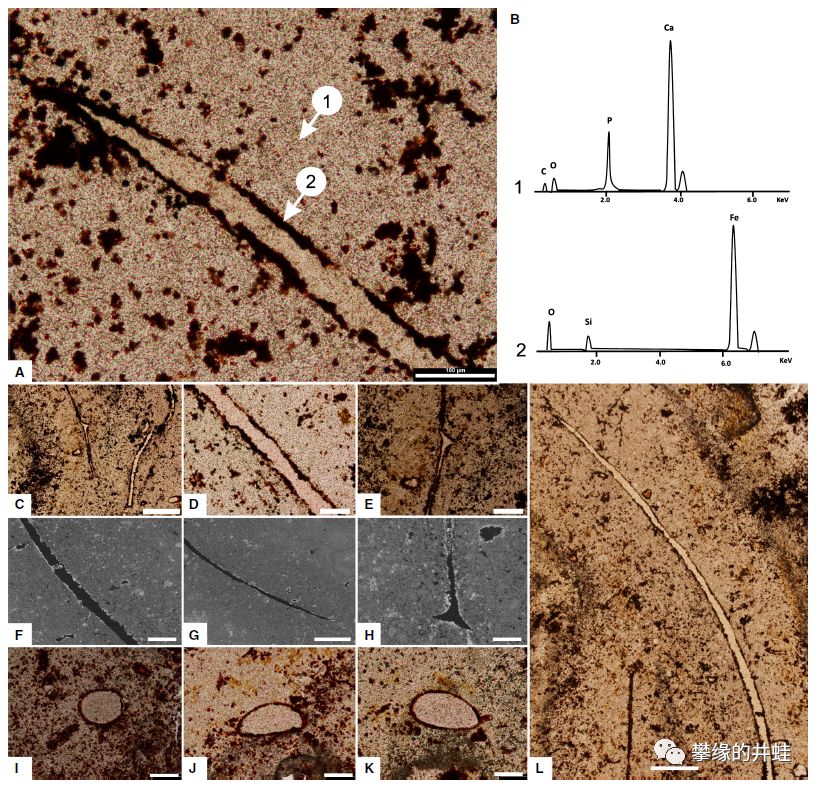
目前发现的最早的疑似毛发化石出现在俄罗斯的晚二叠世动物粪便化石中(上图中的纤维状物)。如果它真的是毛发的话,应该是来自被吃掉的猎物,也可能是动物清理自己时吞下的。研究者不能确定这些疑似毛发来自哪种动物,不过和哺乳动物亲缘关系最近的兽头兽和犬齿兽无疑可能性是最大的。图片来源自 [20]。
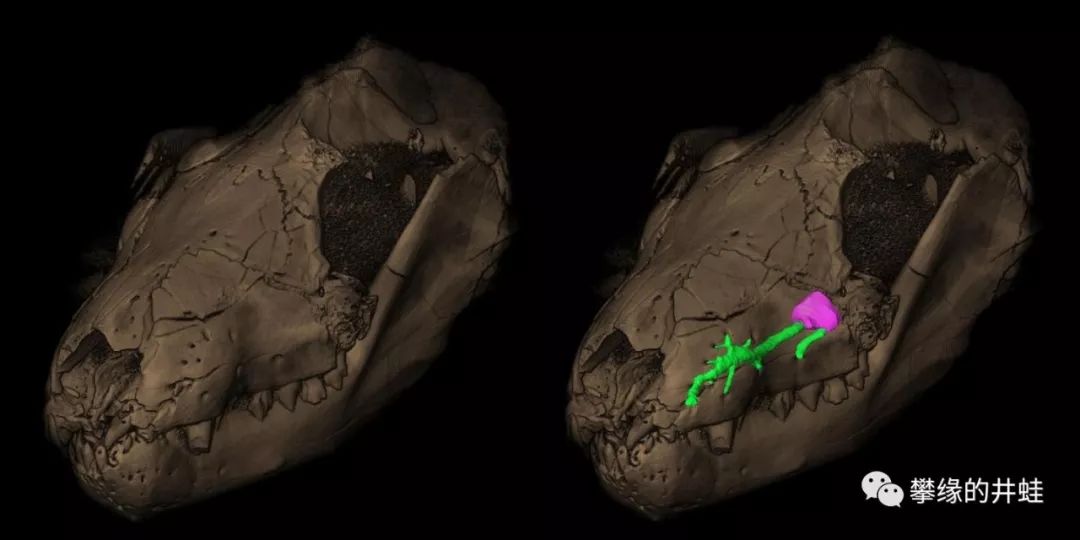
南非发现的早三叠世犬齿兽类:三尖叉齿兽Thrinaxodon头骨化石。注意口鼻部的微小孔洞,复原后发现其中联通有神经纤维(绿色网络)。研究者推测这些小孔可能是胡须生长的位置,用来探知障碍物和感受空气震动。这大概就是毛发最早的功能。图片来源自[21]。
除了感官之外,早期兽孔类也可能用毛发来保暖(兽头亚目的 Glanosuchus 就可能是恒温动物的),或者硬化毛发成豪猪那样的尖刺用来防卫。体表生长起厚密的毛发后,防水,抗菌,除霉就成了大问题。兽孔类可能因此演化出发达的皮下腺体,分泌油脂和抗菌化合物来保养毛皮。经过一连串的变异后,一种变性的分泌物最终成为它们哺育下一代的专用食品。
Q2:为什么复原图上的兽孔目动物都没有耳朵呢?
A2:前面的回目提到过,哺乳动物的下颌由单独的齿骨构成。其他的骨头有的消失(比如隅骨和上隅骨),有的放到耳朵里去了(关节骨articular)。同时移入耳朵的还有上颌的方骨(quadrate)。改造后的关节骨称为锤骨(malleus),改造后的方骨称为砧骨(incus),它们和原本在耳朵里的镫骨(stapes)组成了一套独特的听小骨(ossicle)系统,可以把鼓膜(eardrum)的震动传高效地送给神经系统。
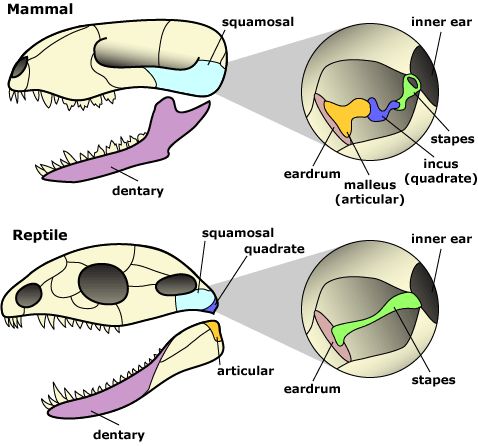
哺乳动物的三块听小骨(上)和其他羊膜类的单一镫骨(下)对比。图片来源自网络。
在这种包含三块听小骨的中耳(middle ear )大幅提升了对空气震动的感知能力。在它出现之前,兽孔类可能对地面,树干等固体基材的震动更敏感。
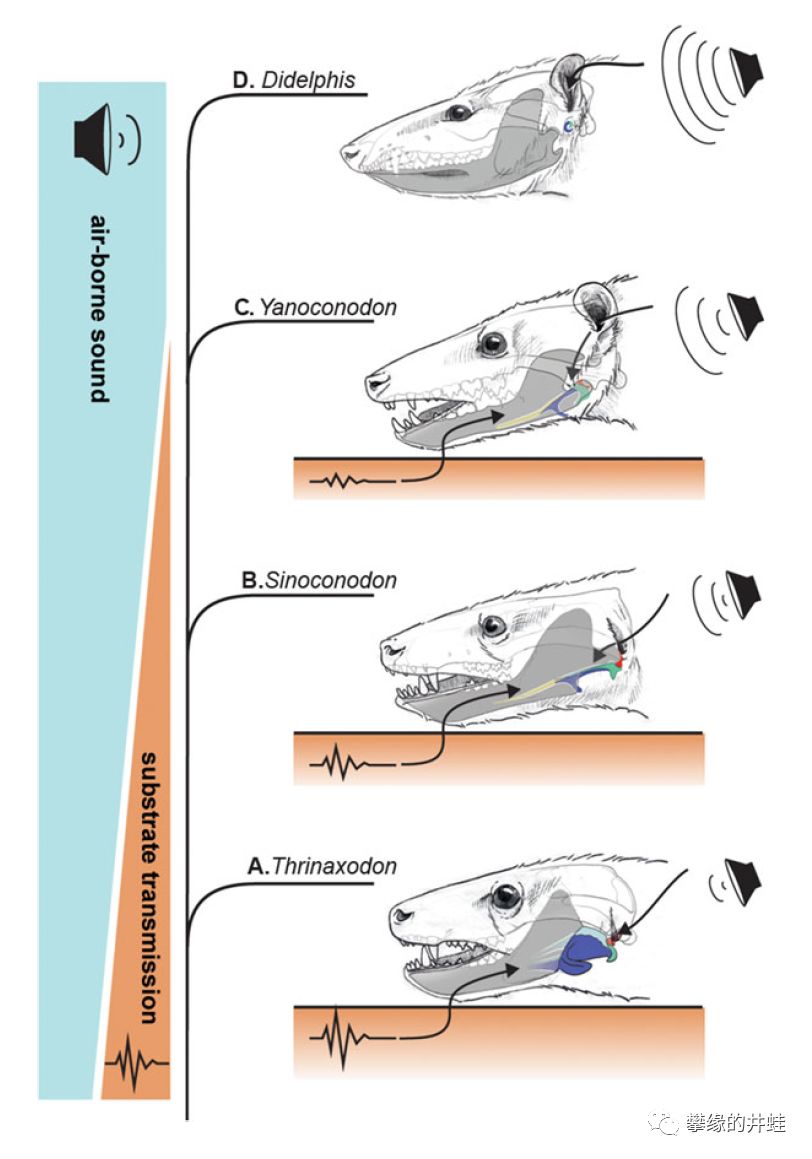
犬齿兽类到哺乳动物的听觉转变示意。A:三尖叉齿兽(犬齿兽类,早三叠世);B:中国锥齿兽(哺乳形类或哺乳动物,早侏罗世);C:阿氏燕兽(哺乳动物,早白垩世);D:现代负鼠。随着听小骨的形成,动物的听觉从主要接收固体震动转为基本完全接收空气震动。图片来源自[22]。
所以在听小骨出现前,兽孔类不大可能出现哺乳动物那样用来收集扩大空气震动的耳朵(外耳,outer ear )。
Q3:兽孔类卵生还是胎生呢?
A3:卵生。
好了,本次回答结束。谢谢!
地球名片
生物分类:动物界-脊索动物门-脊椎动物亚门-羊膜类-合弓纲-兽孔目-兽头亚目
存在时间:二叠纪中期 至 三叠纪中期
现存种类:无
化石种类:未找到完整统计
生活环境:陆地
代表特征:异型齿比丽齿兽发达,但犬后齿的演化程度不如犬齿兽亚目
主要类群:Scylacosauridae科,弱颌兽科,霍夫梅里兽科,Whaitsiidae,包氏兽科
参考文献
[1] Tom S. Kemp, Phylogenetic interrelationships and patternof evolution of the therapsids: testing for polytomy. Palaeont. afr. (December2009), 44: 1–12
[2] FERNANDO ABDALA, REDESCRIPTION OF PLATYCRANIELLUSELEGANS (THERAPSIDA, CYNODONTIA) FROM THE LOWER TRIASSIC OF SOUTH AFRICA, ANDTHE CLADISTIC RELATIONSHIPS OF EUTHERIODONTS. Palaeontology, Vol. 50, Part 3,2007, pp. 591–618
[3] https://www.deviantart.com/
[4] https://www.upi.com/Science_News/2018/06/08/New-saber-tooth-species-help-scientists-trace-evolution-of-mammals/9381528461260/
[5] Adam K. Huttenlocker , Christian A. Sidor, Roger M. H.Smith, A new specimen of Promoschorhynchus (Therapsida: Therocephalia:Akidnognathidae) from the Lower Triassic of South Africa and its implications fortheriodont survivorship across the Permo-Triassic boundary. Journal ofVertebrate Paleontology, 31:2, 405-421, 2011, DOI: 10.1080/02724634.2011.546720
[6] Huttenlocker and Smith, New whaitsioids (Therapsida:Therocephalia) from the Teekloof Formation of South Africa and therocephaliandiversity during the end-Guadalupian extinction. PeerJ 5:e3868, 2017, DOI10.7717/peerj.3868
[7] M. F. Ivakhnenko, The First Whaitsiid (Therocephalia,Theromorpha) from the Terminal Permian of Eastern Europe. PaleontologicalJournal, 2008, Vol. 42, No. 4, pp. 409-413
[8] JULIEN BENOIT, A review of the “venomous therocephalian”hypothesis and how multiple re-portrayals of Euchambersia have influenced itssuccess and vice versa. Bull. Soc. géol. France, t. 187, no 4-5, pp. 217-224,2016
[9] Benoit J, Norton LA, Manger PR, et al., Reappraisal ofthe envenoming capacity of Euchambersia mirabilis (Therapsida, Therocephalia)using μCT-scanning techniques. PLoS ONE 12(2): e0172047, 2017,doi:10.1371/journal.pone.017204
[10] Benoit J, Manger PR, Fernandez V, et al., CranialBosses of Choerosaurus dejageri (Therapsida, Therocephalia): Earliest Evidenceof Cranial Display Structures in Eutheriodonts. PLoS ONE 11(8): e0161457, 2016,doi:10.1371/journal.pone.0161457
[11] Heidi Fourie, Bruce S. Rubidge, The postcranialskeleton of the basal therocephalian Glanosuchus macrops (Scylacosauridae) andcomparison of morphological and phylogenetic trends amongst the Theriodontia.Palaeont. afr. (December 2009) 44: 27–39
[12] T. S. KEMP, The origin of mammalian endothermy: aparadigm for the evolution of complex biological structure. Zoological Journalof the Linnean Society, 147, 473–488, 2006
[13] Yu. A. Suchkova, V. K. Golubev, A New PrimitiveTherocephalian (Theromorpha) from the Middle Permian of Eastern Europe.Paleontological Journal, 2019, Vol. 53, No. 3, pp. 305–314.
[14] Christian A. Sidor, James A. Hopson, Ghost lineages and"mammalness": assessing the temporal pattern of character acquisitionin the Synapsida. Paleobiology, 24(2), pp. 254-273, 1998
[15] J. Botha-Brink, F. Abdala, A new cynodont record fromthe Tropidostoma Assemblage Zone of the Beaufort Group: implications for theearly evolution of cynodonts in South Africa. Palaeont. afr., 43: 1–6, 2008
[16] J. BOTHA, F. ABDALA, R. SMITH, The oldest cynodont: newclues on the origin and early diversification of the Cynodontia. ZoologicalJournal of the Linnean Society, 149, 477–492, 2007
[17] Marc J. Van den Brandt, Fernando Abdala, Cranialmorphology and phylogenetic analysis of Cynosaurus suppostus (Therapsida,Cynodontia) from the upper Permian of the Karoo Basin, South Africa.Palaeontologia africana 52: 201–221
[18] Yu. A. Suchkova, V. K. Golubev, A New PermianTherocephalian (Therocephalia, Theromorpha) from the Sundyr Assemblage ofEastern Europe. Paleontological Journal, Vol. 53, No. 4, pp. 411–417, 2019
[19] Danielle Dhouailly, A new scenario for the evolutionaryorigin of hair, feather, and avian scales. J. Anat., 214, pp 587–606, 2009,doi: 10.1111/j.1469-7580.2008.01041.x
[20] Bajdek, P., Qvarnström, M., Owocki, K., et al., Microbiotaand food residues including possible evidence of pre-mammalian hair in UpperPermian coprolites from Russia. Lethaia, Vol. 49, pp. 455–477, 2016, DOI10.1111/let.12156
[21] http://sciencenewsjournal.com/therapsids-developed-hair-order-survive-millions-years-ago/
[22] Luo ZX., Schultz J.A., Ekdale E.G., (2016) Evolution ofthe Middle and Inner Ears of Mammaliaforms: The Approach to Mammals. In: ClackJ., Fay R., Popper A. (eds) Evolution of the Vertebrate Ear. Springer Handbookof Auditory Research, vol 59. Springer, Cham