

原标题:【地球演义】第二百十八回 寒武纪的余烬:棘皮动物篇(下)
继续古生代棘皮动物的话题。不知道究竟是什么原因,促使我们的这个远亲在出现不久就偏离了两侧对称的身体模式,走上一条只能用魔幻来形容的演化道路。




井蛙真的感觉棘皮动物有一种把魔界的碎片传送到这个世界的能力——特别是当它们聚集在一起的时候。图片来源自网络或见水印。
绝大多数动物都呈现两侧对称的外形,这绝非偶然,因为这样的身体构型最适合自由运动的生活模式。两侧对称使得动物自然而然地形成了“首”-“尾”,“背”-“腹”四个方向,为定向移动奠定了基础。这个框架搭好之后,一切都变得顺理成章:发达的神经和感官系统促使头部形成;贯穿身体的消化道和有序排布的其他内脏;对称排列,协同运作的体壁肌肉和附肢;支撑身体的外壳,贝壳和骨骼,等等等等。拥有这些武器后,两侧对称的动物可以不像植物一样固定在海底,或者做布朗运动般的随波逐流,它们探查,感知,搜索,追逐和逃避,生存竞争的烈度大幅提高,这又催生出更多的结构,更精密的分工和组合,和更强大高效的功能。
上面的演化逻辑在棘皮动物这里彻底失效。失去了两侧对称的身体后,棘皮动物再也没有演化出统筹全身的神经中枢,发达的感觉器官,和强健的肌肉组织,再加上骨板的限制,它们只能慢吞吞地完成一些有限的动作,对外界刺激做一些简单的本能反应。这样的身体条件,自然无法追捕猎物,逃避敌害。只能固着或者缓慢爬行,以过滤海水或者吞食沉积物为生。
一般来说,海水和淤泥中的有机碎屑比较丰富和稳定,以此为食的动物又不大运动,能耗很低,除非特殊情况,否则不至于会饿肚子。而且棘皮动物一身薄皮碎骨,一般的捕食者对这些扎嘴咯牙还没营养的东西没什么兴趣。相比那些辛苦奔波的两侧对称动物,它们的生存压力要小一些。
非两侧对称的身体和简单的内脏系统很可能还带来了一个意想不到的好处。两侧对称的动物如果发生破坏对称性的变异,无论躯干还是肢体,一般都会严重损害感知和运动能力,而被很快淘汰。但棘皮动物的骨板发生错位和变形,体盘发生扭曲和畸形,腕长错了位置,长一点短一点,多一根少一根,或者中间分个岔,好像都不会带来太大的问题。

两侧对称动物的畸形(如左图)会彻底打乱呼吸,神经,消化,感官,运动和生殖系统的精确布局,造成严重后果(节肢动物很可能会蜕皮失败,直接卡死在自己的身体里),很难顺利成长并留下后代。但是,右图中的海星也发生了严重的畸形:本来是五辐对称的身体变成了八辐对称,凭空多了近一倍的体盘和腕,却基本不影响它的生存和繁殖,甚至长得比上方的正常海星更大更粗壮。图片来源自网络。

棘皮动物还有着动物界最强大的再生能力,就算被切碎,也有很大机会长成一群完整的个体。它们没有大脑,心脏,眼睛这样不可再生的精密器官,除非被放到太阳下曝晒,否则不会死于出血,窒息,休克和脏器破裂。没有哪些部分不可以失去,也没有哪些部分不可以长回来。图片来源见水印。
井蛙强烈怀疑(无力查证,就只能怀疑):正是这种强大的容错能力,使得棘皮动物得以在各种奇葩的道路上深入探索。从寒武纪到奥陶纪,棘皮动物的多样性经历了一轮爆发式增长。就像被施了魔法一样,各种奇形怪状的骨板,刺棘,体盘和须腕以各种诡异的方式组合起来,成为古生代海洋中的一道奇景。
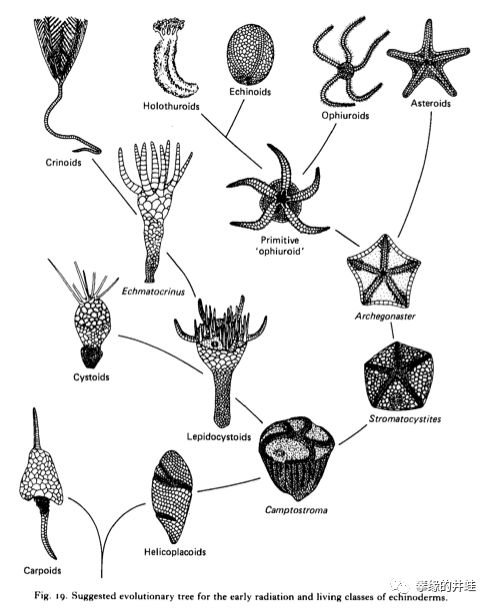
关于棘皮动物的演化过程有众多理论。上图只是示意了其中的一种。基本上,争论集中在辐射对称和五辐对称,腕和茎的分化,步带沟和水管系统这些特征出现的先后顺序和发展历程。图片来源自网络。

即使是那些外表看起来完全不同的棘皮动物类群,比如海座星纲(左),海旋板纲(中)和始海百合纲(右),也能够找到同源的结构和器官(红色部分:步带沟;蓝色和绿色部分:体盘;黄色部分:茎和固着器),只不过变形得太厉害了。图片来源自[1]。
如果说海扁果类反映了棘皮动物的身体怎样从两侧对称逐渐过渡到不对,那么另一些类群则展示了辐射对称的出现过程:
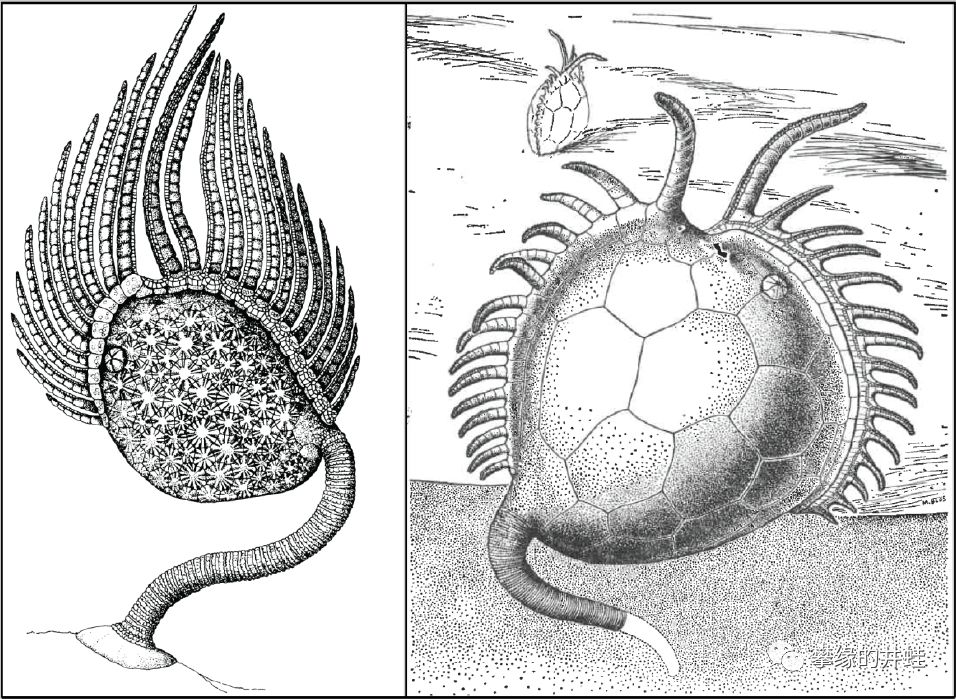
两种拟海百合纲动物(左:Amygdalocystites;右:Platycystites)的复原图。它们的样子介于海扁果类和有柄类(海蕾亚门,海百合亚门)之间。一根远离口部的“腕”变得特别发达,用来支撑和固定身体,这就是“茎”的雏形。它们不再平趴,而是竖立在海底。体盘不再扁平,开始变得浑圆,向着“萼”的方向发展。图片来源自[2]。

一些拟海百合纲动物的萼部化石。它们已经产生了明显的适应直立滤食的特征,比如位于同侧的口和肛门,意味着消化道发生了180度的扭转(就像帚虫,苔藓虫和腕足动物那样)。腕开始出现特殊的排布方式,这样有助于击打水流和传送食物。图片来源自[2]。
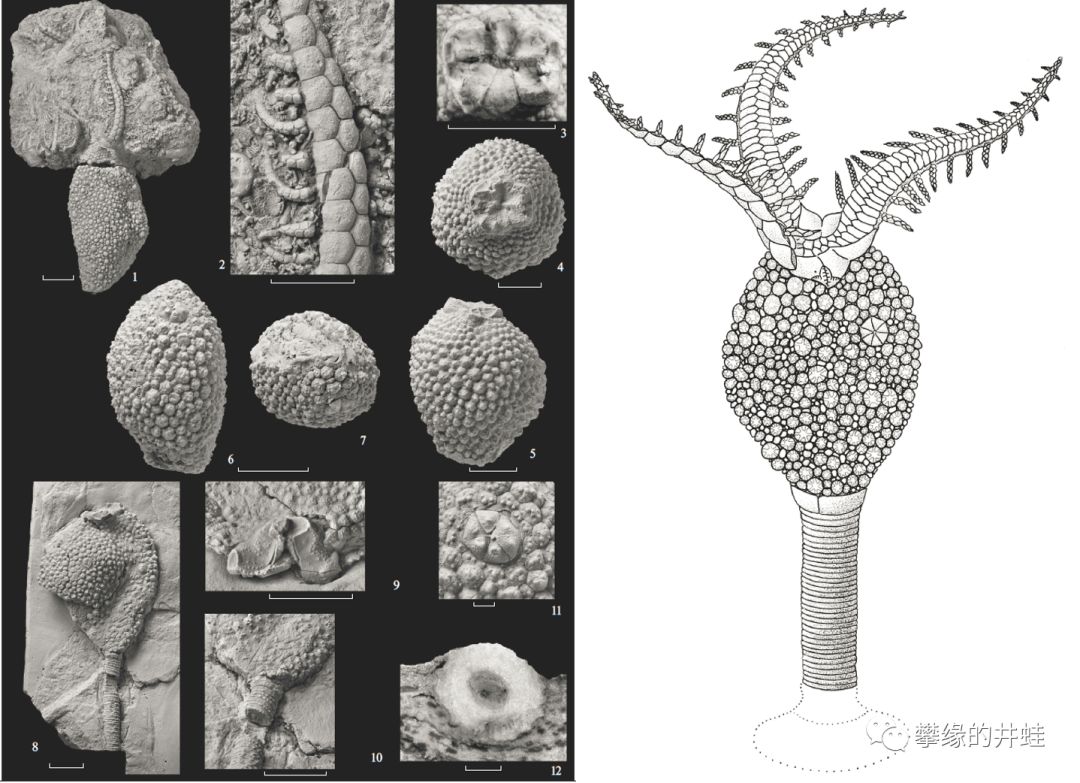
带有腕的拟海百合化石很少,上图是爱沙尼亚发现的晚奥陶世拟海百合Achradocystites schmidti化石和复原图。它的三根腕已经接近辐射对称,而且腕上开始出现分枝。开始向着海百合的形态发展。图片来源自[3]。
就这样,棘皮动物终于和刺胞动物殊途同归,走上了辐射对称的道路。对于固着或半固着滤食的动物来说,这确实是最方便的体型,并在不同的无脊椎动物类群中反复出现:比如苔藓动物门,环节动物的缨鳃虫,节肢动物的蔓足纲,半索动物的羽鳃纲和笔石纲等等。

捷克发现的寒武纪晚期始海百合Akadocrinus jani化石。始海百合纲是海蕾亚门的早期演化类型,也是最早演化出辐射对称的身体的棘皮动物之一。图片来源自[4]。

两类海旋板:Helicoplacidae(左)和Helicocystoidae(右)的化石和复原图。它们的扭旋可以看做是辐射对称的一种变体:骨板不是竖直排列,而是扭转盘旋。图片来源自[1]。

海旋板没有腕,不能主动捞取食物,身边的水流情况至关重要。研究者模拟了两种海旋板的形态对流经自身的水流的影响。结论是它们也会在口附近制造缓慢的湍流,延长食物碎屑的停留时间,提高进食的效率。图片来源自[1]。
海旋板纲只在寒武纪中期昙花一现,始海百合纲也在志留纪灭绝。继之而起的是海蕾亚门的其他类群,和海百合亚门。它们把辐射对称发展到了顶峰,都演化出动物界最像植物的体态,规整的五辐对称结构,灵活的长茎,成群的羽腕(这个部分也叫冠),还有令人演化缭乱进食动作。它们在海底成片生长,就像恶魔精心打理的花田。

海蕾萼部的骨板高度愈合,变成一个坚固而精致的小球,在石化后还能保持完整的立体外形,是古生物爱好者和化石玩家们的至爱。图片来源自[5]。

一些奥陶纪到二叠纪的海蕾萼部化石。海蕾纲在古生代晚期极为繁盛,演化出许多种类,但基本特征没有太多改变。图片来源自[5]。

大多数海蕾化石都是茎和腕脱落后剩下的光溜溜的萼部。这个标本难得地保留了腕和一部分茎,让我们看到这种动物或者时是什么样子。图片来源自网络。

模拟水流通过海蕾萼部的变化情况。像海蕾和海百合这样高度演化的类群,可以用肌肉控制茎的朝向和弯曲角度,最大限度地制造湍流,发达的羽腕主动捞取食物,灵活地送入口中。生存能力自然比海旋板和始海百合更强。图片来源自[6]。
海百合的萼部小,骨板也没有愈合,死后很容易解体,所以化石以零散的碎片居多。但它们的腕远比海蕾粗壮,具有复杂的多重羽状分枝。

上图是一些意大利发现的晚奥陶世海百合各部分的化石,属于不同的种类。A:萼部(中间的圆圈是茎连接的部分);B,C:茎末端的固着器;D,E:带有腕(冠)的萼部。把这些部分结合起来,就可以想象出这类动物活着的样子。图片来源自[7]。
还有一些古生代的海百合简直就是在挑战人类想象力的极限,实在难以相信地球上居然曾经出现过这样的生物(而且还是我们的近亲)。可偏偏它们留下的化石非常完整,无可抵赖:
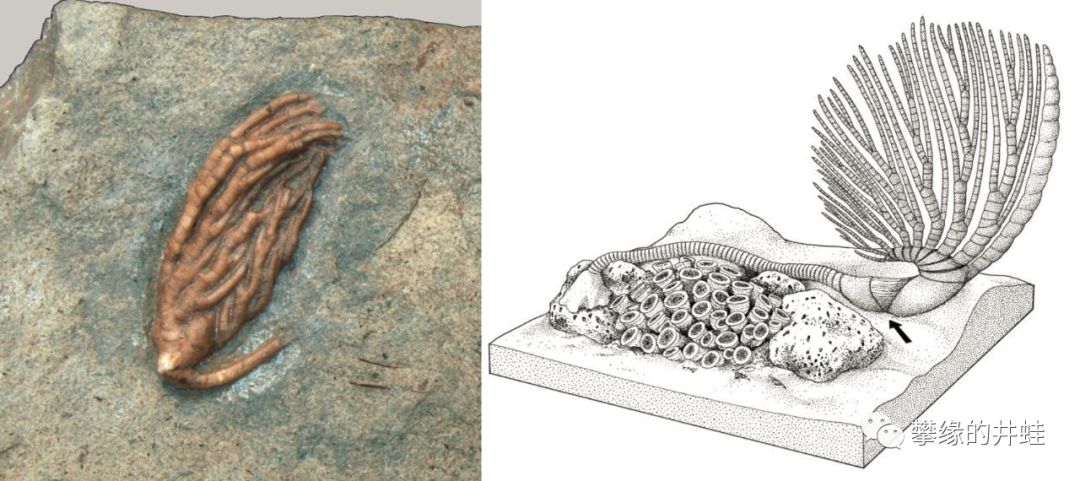
奥陶纪海百合Calceocrinus。图片来源自[8]。

志留纪海百合Dimerocrinites。它的茎末端没有固着器,可以自由移动,盘绕在硬质的基体上。图片来源自[8]。

石炭纪海百合Dorycrinus。图片来源自[8]。

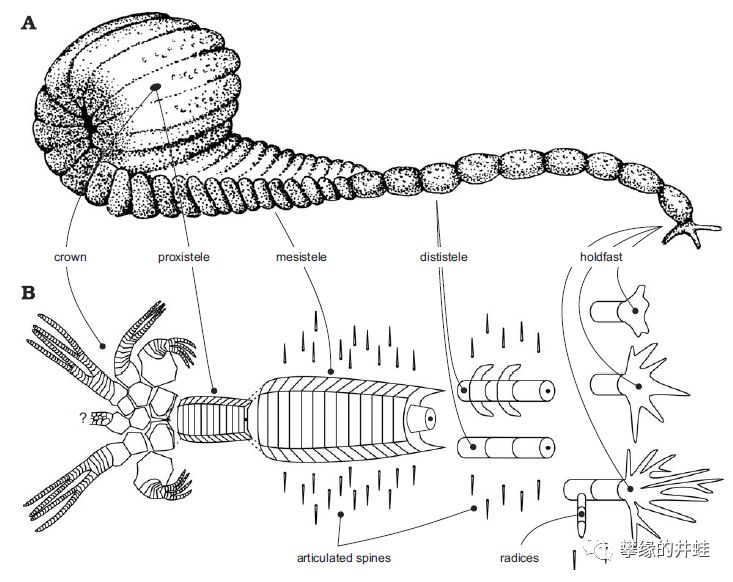
泥盆纪海百合Ammonicrinus。井蛙第一次看到它时,心里只想这是来自地狱的犰狳吗?图片来源自[9]。
当然还有许多形态介于海蕾,海百合甚至海扁果之间的类群,甚至根本不知道归属到哪类的怪咖。

一些晚奥陶世具柄的棘皮动物的萼部化石。它们的茎和腕没有保存,更令人浮想联翩。图片来源自[7]。

海盘囊纲(Cyclocystoidea)是最神秘的一类棘皮动物,出现在中奥陶世到早石炭世,留下的化石是一些扁平的圆盘,由从中心向四周辐射的骨片组成。关于海盘囊的完整形态,内部结构,生活习性和分类归属几乎完全未知。上图是摩洛哥发现的中奥陶世海盘囊化石。图片来源自[10]。
具柄的棘皮动物在古生代极其繁盛。然而魔法终有失效的一天,棘皮动物也不可能超然在生存竞争之外。随着节肢动物,软体动物和脊椎动物的竞争日趋白热化,棘皮动物的地位也日益边缘化。在奥陶纪以后,棘皮动物再也没有演化出新的纲一级的分类单元。而每一次大灭绝,都会把几个原有的纲从地球上轻轻抹去。二叠纪末大灭绝带走的是海座星纲和海蕾纲,海百合纲奄奄一息,而海蛇函纲终于没能撑到下一次大灭绝的到来。

海百合纲多样性(科数)在地质史上的变化情况。它们差一点就被显生宙最大的灭绝事件彻底吞噬。少数残余苦苦熬过接踵而来的又一场灾难后,终于在侏罗纪又迎来了新的繁荣。在中生代的回目里,还有它们的位置。而海蕾就没有这么幸运,它们的身影永远留在了古生代,一同消失的还有和它们有着说不清道不明的暧昧关系的宽角螺(参见第一百三十一回螺宫)。图片来源自网络。

海座星纲是另一个沉没在古生代终点前的棘皮动物类群。不过一些和它们关系密切的近亲在中生代和新生代欣欣向荣。限于篇幅,还是等到写海星亚门和海胆亚门的时候再顺带连海座星一起介绍吧。图片来源自[11]。
生物分类:动物界-棘皮动物门-海蕾亚门
存在时间:寒武纪早期至二叠纪末
现存种类:无
化石种类:不明
生活环境:海洋
代表特征:具萼,羽腕和茎,后期种类萼部骨板愈合,五辐对称明显。萼内体腔不扩展至腕内部。
代表类群:始海百合纲,海蕾纲,海林檎纲,拟海蕾纲
参考文献:
[1] Louis Dudit, Imran Rahman, John Cunnigham, Investigatingthe evolution of symmetry in Cambrian echinoderms.
[2] Limbeck, M.R.Fossil Focus: Paracrinoids. Palaeontology Online, Volume 8, Article 9, 1-7
[3] S. V. Rozhnov, Ordovician Paracrinoids from the Baltic:Key Problems of Comparative Morphology of Pelmatozoan Echinoderms. Paleontol.J. (2017) 51: 643, https://doi.org/10.1134/S0031030117060065
[4] NOHEJLOVÁ M. &FATKA O. (2017). Revision of the BARRANDE's specimen "Tige d'une Cystidée
indéterminée"(Cambrian, Echinodermata, Eocrinoidea). Carnets Geol., Madrid, vol. 17, No. 8,p. 153-
160, DOI: 10.4267/2042/62768
[5] Bauer, J.E. Fossil Focus: Blastoids. PalaeontologyOnline, Volume 8, Article 7, 1-7
[6] Stephen K. Donovan, Gary D. Webster, Johnny A. Waters, Alast peak in diversity: the stalked echinoderms of the Permian of Timor.Geology Today, Vol. 32, No.5, September-October 2016
[7] Colin D. Sumrall, Bradley Deline, Jorge Colmenar, etal., NEW DATA ON LATE ORDOVICIAN (KATIAN) ECHINODERMS FROM SARDINIA, ITALY.Progress in Echinoderm Palaeobiology.
[8] http://www.fossilcrinoids.com/index.html
[9] Bohatý, J. 2011. Revision of the flexible crinoid genusAmmonicrinus and a new hypothesis on its life mode. Acta PalaeontologicaPolonica 56 (3): 615–639
[10] James Sprinkle, Mike Reich, Bertrand Lefebvre, COMPUTEDTOMOGRAPHY (CT) SCANS OF A NEW ORDOVICIAN CYCLOCYSTOID FROM MOROCCO AND ITSORIENTATION AND LIFE MODE. Cuadernos del Museo Geominero, 19. Instituto GeológicoyMinero de España, Madrid. ISBN: 978-84-7840-961-7
[11] COLIN D. SUMRALL, FIRST DEFINITE RECORD OF PERMIANEDRIOASTEROIDS: NEOISOROPHUSELLA MASLENNIKOVI N. SP. FROM THE KUNGURIAN OFNORTHEAST RUSSIA. J. Paleont., 83(6), 2009, pp. 990–993
感兴趣的话,不妨关注攀缘的井蛙,每天琢磨点新东西: